Moyamoya Hastalığı Nedir? Belirtileri ve Tedavi Seçenekleri Nelerdir? için yorumlar kapalı
Moyamoya hastalığı; beyinde bulunan bazı arterlerin (damarların) daralması sonucunda ortaya çıkan kronik, ilerleyici bir serebrovasküler hastalıktır. Bu hastalık, Willis poligonunu oluşturan arterlerin bilateral, ilerleyici darlığı veya tıkanıklığına bağlı olarak gelişen kollateral damarların karakteristik bir özelliği olarak tanımlanır. Bu durum, beyne yeterli kan akışının sağlanamamasına ve oksijenin yetersiz olduğu bölgelerde iskemik semptomlara (kan eksikliğinden kaynaklanan belirtiler) yol açabilir. Hastalığın sebepleri net olarak bilinmemekle birlikte, genetik ve konjenital faktörlerin etkili olduğu düşünülmektedir. Ayrıca, bazı tümöral hastalıklarla birlikte görülebilir.[1], [2], [3], [4]
Belirti ve Bulgular
Moyamoya hastalığının belirti ve semptomları yetişkinler ve çocuklarda farklılık gösterebilir.
Çocuklarda gözlenen iskemik semptomlar şu şekildedir:
Tek taraflı vücut zayıflığı veya felç,
Konuşma güçlükleri,
Nöbet geçirme,
Baş ağrısı,
Yürüme zorluğu.
Erişkinlerde gözlenen iskemi ve kanama semptomları ise şu şekildedir:
Aniden başlayan şiddetli baş ağrısı,
Bilinç kaybı,
Bulantı ve kusma,
Göz kararması veya çift görme.
Yaş ve cinsiyete bağlı semptom farklılıkları ise şöyle sıralanabilir:
Çocuklarda genellikle iskemik semptomlar öne çıkar.
Erişkinlerde ise kanama ve iskemi semptomları eşit ağırlıkta görülür.
Kanama insidansı erişkinlerde %60, çocuklarda %10 olarak bildirilmiştir.
İskemik belirtiler, erişkinlerde genellikle komplet strok, çocuklarda ise geçici iskemik atak şeklinde ortaya çıkar.
İntrakraniyal kanama, genel olarak kötü bir prognostik faktördür.
Çocuklarda moyamoya, inme olgularının %6’sından sorumlu olabilir.
Moyamoya hastalığı çocuklarda kötü bir prognoza sahip olabilir, özellikle semptomlar 2 yaşından önce başladıysa bu beklenen bir durumdur.
Bu belirtiler, hastalığın seyri ve yaşa bağlı olarak değişebilir; bu nedenle bireysel durumlar dikkate alınmalıdır.[1], [2], [4], [5], [6]
Hastalıkla İlişkili Genler, Etken Faktörler ve Risk Faktörleri
Moyamoya hastalığı, genetik ve çevresel faktörlerin karmaşık etkileşimi sonucu ortaya çıkan bir nörovasküler bozukluktur. Genetik yatkınlık, izole moyamoya hastalığı için poligenik bir etiyoloji öne sürmektedir. Bu durum, birden fazla genin hastalığın gelişiminde rol oynamasına işaret etmektedir. Bilinen genetik mutasyonlar arasında BRCC3, MTCP1 ve GUCY1A3 genlerindeki değişiklikler öne çıkmaktadır.[5]
Genetik araştırmalar ve moleküler çalışmalar, moyamoya hastalığının temelinde yatan genetik faktörleri anlamaya yöneliktir. Ancak, hastalığın tam anlamıyla çözümlenmesi ve etkili tedavi stratejilerinin geliştirilmesi için daha fazla çalışma gerekmektedir.[5]
Teşhis Yöntemleri
Moyamoya hastalığının teşhisi, nadir görülmesi ve bariz olmayan belirti ve semptomlardan dolayı zor olabilir. Teşhis, klinik bulgular ve görüntüleme bulguları temelinde şüphelenilerek konur. Teşhisin doğrulanması için standart yöntem serebral anjiyografi olarak kabul edilir.[7]
Moyamoya hastalığının T1 ağırlıklı MR görüntüsü. Bazal ganglionda görülen akım boşluğu ok ile işaretlenmiştir.
Serebral anjiyografi, moyamoya hastalığını doğrulamak için kullanılan standart bir yöntemdir. İşlem sırasında, kontrast madde enjeksiyonu ile beyin damarlarının röntgen filmi çekilir. Ancak, bu yöntem intravenöz kontrast kullanımına bağlı alerjik reaksiyonlar ve nadiren kontrast maddeye bağlı böbrek hasarı gibi riskleri içerebilir.[7]
Manyetik Rezonans Anjiyografi (MRA), serebral anjiyografinin alternatifi olarak kullanılabilir. Bu yöntem, manyetik rezonans görüntüleme (MRG) prensiplerini kullanarak damarların görüntülerini oluşturur. Genellikle serebral anjiyografiye göre daha düşük risk içerir.[7]
Tanıda etkili olan bir diğer unsur, hastanın klinik belirtileri ve görüntüleme bulgularıdır. Moyamoya hastalığının belirgin bulguları genellikle supraclinoid iç karotis arterleri ve bunların proksimal dallarını içerir.[7
Moyamoya hastalığının teşhisinde serebral anjiyografi genellikle en güvenilir yöntem olmakla birlikte, MRA gibi non-invaziv yöntemler de kullanılabilir. Teşhis sürecinde hastanın genel sağlık durumu, alerji öyküsü ve kontrast madde kullanımına karşı hassasiyet dikkate alınmalıdır. Teşhis, uzman bir sağlık profesyoneli tarafından detaylı bir değerlendirme sonucunda konmalıdır.[7]
Moyamoya hastalığı olan bir hastanın sağ iç karotid arterini (ICA) gösteren bir DSA (Dijital Substraksiyon Anjiyografi) görüntüsüdür.
Tedaviler veya İdare Yöntemleri
Moyamoya hastalığının ilerlemesini durdurmak veya intrakraniyal arteriyopatiyi tersine çevirmek için spesifik bir tedavi mevcut değildir.[7] Tedavi, hastalık komplikasyonlarına karşı önlem almak, semptomları hafifletmek ve bilişsel bozulmayı önlemek amacıyla kullanılan ilaç tedavisini içerir. Atak sonrası fiziksel, mesleki ve konuşmaya yönelik terapiler de gerekebilir.[6], [7]
İlaç tedavisi, hastalık komplikasyonlarını önlemek amacıyla kullanılır. Bu ilaçlar genellikle kan pıhtılaşmasını önleyen antikoagülanlar, kan basıncını düzenleyen ilaçlar ve antiplatelet ajanları içerir. Ancak, ilaçların kullanımı bireyin genel sağlık durumu ve diğer ilaçlarla etkileşimlere bağlı olarak riskler içerebilir.[6], [7]
Fiziksel, mesleki ve konuşmaya yönelik terapiler ise atak sonrası fonksiyonel yetenekleri restore etmek ve geliştirmek amacıyla uygulanır. Fizik tedavi, mesleki terapi ve konuşma terapisi, hastanın ihtiyacına bağlı olarak uygulanır ve hastanın yaşam kalitesini artırmada önemli bir rol oynar.[6], [7]
Ek olarak cerrahi revaskülarizasyon prosedürleri, beyine kan akışını restore etmek için kullanılır. Bu prosedürler doğrudan, dolaylı veya kombinasyonlu teknikleri içerebilir. İşlem sırasında olası komplikasyonlar göz önünde bulundurulmalıdır. Cerrahi müdahalenin endikasyonları ve zamanlaması konusunda tartışmalar devam etmektedir. İşlem, iskemik inme riskini azaltabilir ve bilişsel fonksiyonları koruyabilir, ancak belirsizlikler bulunmaktadır.[6], [7]
Moyamoya hastalığının yönetimi multidisipliner bir yaklaşım gerektirir ve bireyin özel durumuna bağlı olarak tedavi planı oluşturulmalıdır. Cerrahi müdahale, belirli durumlarda faydalı olabilir ancak riskler ve faydalar dikkatlice değerlendirilmelidir.
Müddet Tahminleri (Prognoz)
Moyamoya hastalığının seyri, erken teşhis ve uygun tedavi ile iyileştirilebilir. Hastalığın şiddeti, prognozu etkileyen temel bir faktördür. Erken teşhis ve tedavi alan hastalarda genellikle daha olumlu bir seyir görülür. Mortalite oranları yetişkinlerde yaklaşık %5, çocuklarda ise %2 civarındadır; başlıca ölüm nedeni ise hemorajidir. Uzun vadeli sonuçlar hastalığın seyrine, tedaviye yanıta ve zamanında müdahaleye bağlı olarak değişebilir.[7]
Görülme Sıklığı ve Dağılımı (Epidemiyoloji)
Moyamoya hastalığı Asya popülasyonlarında, özellikle Japonya ve Çin’de daha sık görülür. Japonya’da yıllık moyamoya insidansı 0.35/100000, prevalansı ise 3.16/100000’dir. Avrupa’da görülme sıklığı Japonya’nın 1/10’u kadardır. Doğu Asya ülkelerinde yüksek insidans ve prevalans gözlenirken, Batı ülkelerinde bu oranlar daha düşüktür. Afrikalı ve Afrika kökenli Amerikalılarda moyamoya ile ilgili sınırlı veri bulunmaktadır, ancak yapılan çalışmalar bazı etnik gruplarda daha yüksek insidans olduğunu göstermektedir.[5]
Önlem Yöntemleri
Moyamoya hastalığına yönelik kesin bir önlem yöntemi bulunmamakla birlikte, belirli tedavi ve yönetim stratejileri ile hastalığın ilerlemesini kontrol etmek ve komplikasyonları önlemek olasıdır.
Etimoloji
Japonca’da “havada dağılan sigara dumanı” anlamına gelen ve “moyamoya” şeklinde telaffuz edilen “もやもや” terimi, hastalığın anjiyografik görünümünden esinlenmiştir.[1], [2], [5], [8]
^ abcdeMayo Clinic. Moyamoya Disease – Symptoms And Causes. (18 Nisan 2023). Alındığı Tarih: 14 Ocak 2024. Alındığı Yer: Mayo Clinic | Arşiv Bağlantısı
Beyin Kanaması Nedir? Beyin Kanaması Geçirdiğinizi Nasıl Anlarsınız? için yorumlar kapalı
Beyin kanaması, beyin parankiminde oluşan kanamadır ve iskemik inmeden sonra en sık görülen ikinci inme (felç) türüdür. En sık görülen nedenler arasında yüksek tansiyon, serebral amiloid anjiyopati, vasküler malformasyonlar ve antikoagülan tedaviler yer alır. Kanamanın yeri ve derecesine bağlı olarak farklı belirti ve bulgular ortaya çıkar. Semptomlar genellikle bilinçte azalma, uzuvlarda kuvvet kaybı, konuşma bozuklukları ve baş ağrılarıdır.
Klinik olarak beyin kanaması iskemik inmeden güvenilir bir şekilde ayırt edilemez, bu nedenle bilgisayarlı beyin tomografi görüntülemesinin mümkün olan en kısa sürede yapılması gerekir. Akut tedavinin bir parçası olarak sık sık artan kan basıncı ve her türlü kanama diyatezi tedavi edilmelidir. Kafa içi basıncı yükselirse, kafa içi basıncını düşürmeye yönelik önlemler alınmalıdır. Hematomun (kanın damar dışına sızması ve dokular arası boşlukta birikmesi durumu) beyin cerrahisi yoluyla boşaltılması kimi zaman gerekli olabilir.
Belirti ve Bulgular
Hastalığın belirti ve semptomları kanamanın oluştuğu yere göre değişmektedir. Semptomlar genel hatlarıyla şu şekilde sıralanabilir:
Bazal ganglion
Kontralateral hemiparezi (Kanamanın olduğu yerin çapraz tarafında felç)
Lezyonun olduğu yöne gözlerin istemsiz kayması
Afazi (Söz yitimi, beynin konuşma ve anlama bölgelerinde meydana gelen işlev bozukluğu sonucu ortaya çıkan konuşma, konuşulanı anlama, tekrarlama, okuma-yazma gibi becerilerin gerçekleştirilememesi sorunu)
Görme kaybı
Talamus
Bilinç kaybı
Kontralateral hemihipestezi ( Kanamanın olduğu yerin çapraz tarafında duyu kaybı)
Hastalıkla İlişkili Genler, Etken Faktörler ve Risk Faktörleri
İntraserebral kanama riskinin üçte biri genetik çeşitlilikle açıklanabilir; bu nedenle popülasyonlar arasındaki genetik farklılıklar, görülme sıklığındaki farkı kısmen açıklayabilir. Etnik kökenler arasında spontan intraserebral kanama görülme sıklığında da farklılıklar mevcuttur. Bu varyasyonun yaklaşık %42’si etnik köken tarafından açıklanmaktadır. APOE (birden fazla etnik kökende), PMF1/SLC25A44 (Avrupa’da), ACE (Asya’da), MTHFR (birden fazla etnik kökende), TRHDE (Avrupa’da) ve COL4A2 (Avrupa’da) varyasyonları beyin kanaması için en ikna edici şekilde ilişkili olan genlerdir.
Teşhis Yöntemleri
İntraserebral kanama, hayati tehlikesi bulunan tıbbi bir acil durumdur. Bu nedenle zamanında teşhis kritik öneme sahiptir. İntraserebral kanamayı iskemik inmeden güvenilir bir şekilde ayırmak için beyin görüntülemesi yapılmalıdır. Bilgisayarlı Tomografi (BT) ve Anjiyografi veya Magnet Rezonans Tomografi (MRT) ve Anjiyografi başlangıç için uygundur. Acil durumlarda Bilgisayarlı Tomografi, hız ve maliyet bakımından daha kullanışlı olup tanılama becerisi açısından ise Magnet Rezonans Tomografi ile anlamlı bir farkı bulunmamaktadır.
Beyin tomografisinde gözüken bir beyin kanaması
Beyin kanamaları için en büyük risklerden biri yüksek tansiyon hastalığıdır. Yüksek tansiyon zamanla beyin damarlarının iç çeperinin elastik yapısını zedeleyip en sonunda direncini kaybetmesine neden olmaktadır. Bu direnç kaybı da zedelenmenin olduğu damar bölgesinden kanamanın başlamasına yol açmaktadır. Bilgisayarlı Tomografi ile kanamanın tespiti sonrası bu kez kontrast madde ile beyin damarlarının görüntülenmesi, kanamanın kaynaklandığı damarların tespiti ve tedavi stratejisi açısından hekimlere bilgi vermektedir.
Tedaviler veya İdare Yöntemleri
Tedavi yaklaşımları konservatif (koruyucu) ve cerrahi yöntemler olarak ikiye ayrılır. Beyin kanamasının Bilgisayarlı Tomografi’de tanılanması kanamanın zamansal boyunu göstermez. Kanamanın hâlâ devam edip etmediği, sızma şeklinde mi yoksa dramatik şekilde yoğun bir kanama mı olduğu bu görüntülemeyle anlaşılmaz ancak pratik olarak kanamanın devam ettiği varsayılarak bu durumu engelleyici hemostatik stratejiler ilk aşamada uygulanır.
Tüm Reklamları Kapat
Bu aşamada en önemli husus, hastanın ilaç kullanım öyküsüdür. Eğer hasta antikoagülan bir ilaç kullanmıyorsa traneksamik asit koruyucu olarak başlanır. Traneksamik asit, kanın pıhtılaşmasını sağlayan fibrin proteinin enzimatik yıkımını (fibrinoliz) sağlayan plazmin enziminin plazminojen proenziminden oluşmasını tetikler.
Fibrinoliz
Burada ayrı bir parantez, halk arasında kan sulandırıcı olarak da bilinen antikoagülan ilaçlar için açılmalıdır. Kanın damarlardaki akışkanlığını belirleyen bazı faktörler bulunmaktadır. Kimi faktörler kanı daha akışkan yaparken (antikoagülan) kimisi daha viskoz, yani pıhtılı (koagülan) yapmaktadır. Normalde bu iki tip faktör grubu arasında bir denge bulunmaktadır ve böylece dokularımız, kanda bulunan besin ve oksijene herhangi bir gecikme olmaksızın erişmektedir. Kısaca herhangi bir kanamada, yani kanın damarlardan dışarıya doğru aktığı durumlarda koagülan faktörler devreye girip en hızlı şekilde bu sızıntıyı kanı bir miktar pıhtılaştırarak durdurur. Böylece kan kaybı daha az olur ve kan hacmi korunmuş olur. Yine de bu pıhtılaşma sonsuza kadar sürmez, ilgili damar kısmı onarıldıktan sonra bu kez antikoagülan faktörler devreye girer ve kanı tekrar akışkan hale getirir.
Bir sebeple bu denge kanın viskoz olması yönünde bozulmuşsa (kalp ritim bozukluğu, tespit edilmiş bir damar tıkanıklığı veya darlığı, kalp kapağı ameliyatları sonrası, genetik damar onarım faktör eksiklikleri gibi) genellikle hekimler dengeyi tekrar sabitleyebilmek için antikoagülan ilaçlara başlar. Bu ilaçların istenmeyen bir etki olarak dengeyi bu kez de kanın akışkan yöne kaydırması ve en son hali itibarıyla sadece beyin için değil, vücudun herhangi bir yerinde kanama riski açığa çıkarması büyük bir risk faktörüdür.
Eğer beyin kanaması tanısıyla servise alınmış bir hastanın antikoagülan ilaç kullanım öyküsü varsa antidot olarak ilgili ilacın etki mekanizmasına göre bu durumu antagonize etmek gerekir [Vitamin K Antagonisti olan Cumarin veya Warfarin gibi ilaçlara karşı K-Vitamini veya Protrombin konsantreleri (PPSB); Direkt oral Antikoagülan olan Apixaban ve Rivaroxabana karşı Andaxanet alfa; Dabigatrana karşı Idarucizumab ve Edoxabana karşı yine Protrombin konsantreleri].
Beyin kanamasının tanılanması sonrasında hasta mutlaka yakın olarak gözlenmelidir. Bu gözlenme genellikle monitörizasyon ile inme ünitelerinde (Stroke-Unit) veya yoğun bakım ünitelerinde yapılmaktadır. İlk aşamada hastanın yaşamsal fonksiyonlarını gösteren solunum sayısı, kalp atım hızı, vücut sıcaklığı ve düzenli nörolojik fizik muayeneler ile genel sağlık durumu takip edilir. Yine hedef olarak kan basıncının sistolik düzeyde 140 mmHg’nın altında olması hedeflenir. Kanamaya bağlı olarak beyin basıncında artış meydana gelebilir. Bu durumda Mannitol, Gliserin gibi ödem azaltıcı ilaçlar kullanılır.
Cerrahi tedavi yöntemler, karar vermesi çok kolay olmayan, karmaşık karar süreçlerini kapsayan ve hastadan hastaya değişen bir kısım uygulamalara sahiptir. Karar vermede dikkat edilmesi gereken hususlar şu şekilde sıralanabilir:
Kanamanın yeri ve boyutu,
Ventriküler kollaps varlığı,
Hastanın bilişsel durumu,
Yaş,
Eşlik eden hastalıklar.
Beyin kanamasında tercih edilen cerrahi yöntemler genel hatlarıyla şu şekilde sıralanabilir:
Kraniyotomi: Beyni ortaya çıkarmak ve beyin içinde bir prosedür gerçekleştirmek için kafatasının bir kısmının geçici olarak çıkarıldığı cerrahi bir prosedürdür.
Storeotaktik Hematom Aspirasyonu: Trepanasyon (kafatasının cerrahi olarak açılması) ve dura ponksiyonundan sonra kanama boşluğuna bir kateter yerleştirilerek hematomun sıvı kısmı aspire edilir. Daha sonra küçük miktarlarda ancak yüksek etkili antikoagülan madde (r-TPA veya ürokinaz) kanama boşluğuna verilir ve hematom, negatif basınç kullanılarak sıvılaştıktan sonra kanama kalıntısı çıkarılır. Gerekirse prosedür tekrarlanır (birkaç kez).
Endoskopik Hematom Boşaltımı: Trepanasyon ve dural ponksiyon sonrasında kanama boşluğu ultrason kullanılarak endoskopla delinerek temizlenir.
Cerrahi işlemlerden hangisinin tercih edileceği vakanın oluştuğu yer, hastanın hastalıktan ne kadar etkilendiği, cerrahın tecrübesi gibi birçok faktöre bağlıdır.
Müddet Tahminleri (Prognoz)
Beyin kanamaları yüksek ölümcüllük tehlikesi taşımaktadır. İlk 1 ay içinde mortalite oranı %40, ilk bir yıl içinde ise yaklaşık %60 dolaylarındadır. Morbidite oranı ise %40’lara varmaktadır. Tekrarlama riski ilk bir yılda yaklaşık %2, sonraki yıllarda yaklaşık %1 olarak tespit edilmiştir. Prognostik açıdan olumsuz faktörler ise şu şekilde sıralanabilir:
Büyük kan hacmi,
İlk 24 saatte hematomda artış,
Yüksek yaş,
Antikoagülan tedavi veya hepatopati sırasında kanama,
İlişkili intraventriküler kanama,
Obstrüktif hidrosefali.
Görülme Sıklığı ve Dağılımı (Epidemiyoloji)
Beyin kanaması halk arasında inme olarak bilinen hastalığın ikinci en sık sebebidir. İnmenin en sık sebebi, beyin dokusunun beyine kan ulaştıran damarların bir pıhtı sebebiyle tıkanması veya damar çeperinin parçalanması sonucu oluşan disseksiyon sebebiyle beslenemediği iskemik durumlardır. İskemik inme hemorajik inmelere kıyasla yaklaşık 4 kat fazla görülmektedir. Dünyada yaklaşık her 100 bin kişiden 25’inde görülmektedir. Erkeklerde görülme sıklığı kadınlara göre 1,5 kat daha fazladır. Her yaşta görülme ihtimali olmakla beraber, yaş arttıkça görülme sıklığı da artmaktadır.
Önlem Yöntemleri
Önlem yöntemleri risk faktörlerinin azaltılması stratejisine dayanmaktadır. Beyin kanamasının önlem yöntemleri şu şekilde sıralanabilir:
Sigara kullanımının azaltılması,
Alkol tüketiminin kontrolü,
Kan basıncının normalde 140/90 mm Hg’dan az, diyabet veya kronik kalp hastalarında 130/80 mmHg’dan az olması,
Uyuşturuculardan kaçınılması.
Etimoloji
Eski Türkçe méyi veya méŋi “beyin, ilik” sözcüğü ile eş kökenlidir. Bu sözcük Eski Türkçe yazılı örneği bulunmayan méñi biçiminden evrilmiştir. Kan sözcüğü ise eski Türkçe aynı anlama gelen kān sözcüğünden evrilmiştir.
Latince intra, “içeride, içinde, içine” edatından gelen “içeride, içeride, içeride” anlamına gelir ve genellikle ekstra-‘ya karşıdır ve birbirleriyle karşılaştırır. Klasik Latincede intra’nın önek olarak kullanılması nadirdir. Cranium kelimesi Yunanca kranion “kafatası”ndan gelir. Hemorrhage Latince hemorajiden, Yunanca haimorrhagia’dan, haima “kan”dan + rhagē “kırılma, boşluk, yarık”tan gelir.
Araştırmacılar 3000 Yaşındaki Mısır Timsahının Son Yemeğini Ortaya Çıkardı! için yorumlar kapalı
Antik Mısır mumyalarını düşündüğünüzde aklınıza ne geliyor? Belki zihniniz sizi bir okul gezisine, müzede cam bir kutunun içindeki mumyalanmış bir insanla yüz yüze geldiğiniz ana götürüyor. Ya da belki de Hollywood’un tasvir ettiği gibi, kumlu mezarlarından zombi benzeri bir şekilde çıkan, kirli sargıları rüzgârda dalgalanan mumyaları düşünüyorsunuz.
Yakın zamanda yapılan bir çalışmada, Dr. Stephanie Woolham ve ekibi, antik Mısırlı mumyacılar tarafından mumyalanan bir timsahın yaşamının son saatlerine dair olağanüstü detaylar ortaya çıkardı. Bilgisayarlı tomografi (CT) tarayıcısı kullanan araştırmacılar, hayvanın nasıl öldüğünü ve ölümden sonra vücuda nasıl işlem yapıldığını belirleyebildi.
Mısırlılar hayvanları, dünyevi ve ilahi âlemler arasında hareket eden önemli bir dini figür olarak düşünüyordu: Şahinler, gökyüzünde yüksekte uçtukları için güneşe (ve dolayısıyla tanrının kendisine) daha yakın oldukları için güneş tanrısı Horus ile ilişkilendiriliyordu. Kediler ise cesur ve vahşice koruyucu bir anne figürü olan tanrıça Bastet ile bağlantılıydı. Çoğu hayvan mumyası ise adak veya hediye olarak hazırlanıyordu. Hayvan mumyaları, MÖ 750 ile MS 250 yılları arasındaki dönemde doğal dünyanın adeta bir fotoğrafını çekse de bu mumyalanmış türlerin bazıları artık Mısır’da bulunmuyor. Örneğin, antik Mısırlılar her gün Nil kıyılarında kutsal ibisleri (uzun bacaklı, kıvrık gagalı su kuşları) görebiliyorlardı. Bu kuşlar, bilgelik ve yazı tanrısı Thoth’a adak olarak milyonlarca kez mumyalanmıştı. Ancak iklim değişikliği ve çölleşmenin etkileri nedeniyle bu kuşlar artık Etiyopya’ya göç etmiş durumda.
Sıkça mumyalanan bir diğer hayvan da timsahtı. Antik dönemde timsahlar Nil’de yaşasa da 1970 yılında Aswan Barajı’nın tamamlanması onların kuzeyde bulunan Aşağı Mısır’daki deltaya doğru hareket etmelerini engelledi.
Timsahlar, Nil’in Efendisi tanrı Sobek ve tarım arazilerine su ve besin açısından zengin mil sağlayan yıllık Nil taşkını ile ilişkilendiriliyordu.
Timsahlar, tanrı Sobek’e adak olarak muazzam miktarlarda mumyalanıyordu. Firavunlar döneminde Mısır’da kötülükten korunmak için tılsım olarak ya timsah derileri giysi olarak giyiliyor ya da evlerin kapılarının üzerine timsah asılıyordu.Timsah mumyalarının çoğu küçük hayvanlardan oluşuyor, bu da Mısırlıların yumurtaları kuluçkaya yatırma ve yavruları ihtiyaç duyulana kadar canlı tutma imkânına sahip olduklarını düşündürüyor. Arkeolojik kanıtlar, yumurtaların kuluçkaya yatırılması ve yavruların büyütülmesi için ayrılmış alanların keşfi ile bunu destekliyor. Bazı timsahlar ise kutsal hayvanlar olarak görüldüğünden özel ilgiyle bakılıyor ve doğal olarak ölümleri gerçekleşene kadar yaşamalarına izin veriliyordu.
Timsahlar büyüdükçe, timsah bakıcıları için risk arttığından araştırmacılar, daha büyük örneklerin doğadan yakalanıp hızlıca öldürülerek mumyalandığını düşünüyor. Daha büyük hayvanların mumyalanmış kalıntıları üzerinde yapılan araştırmalar, muhtemelen hayvanları hareketsiz hale getirmek ve öldürmek için insanların başlarına darbe vurduğunu gösteriyor. Dr. Stephanie Woolham şöyle diyor:
Birleşik Krallık’taki Birmingham Müzesi ve Sanat Galerisi koleksiyonunda bulunan bir timsah mumyası üzerindeki çalışmalar, bu hayvanların nasıl yakalanmış olabileceğine dair önemli bilgiler veriyor. Mayıs 2016’da, Manchester Üniversitesi’nden birlikte çalıştığım bir araştırmacı ekibinin daha geniş bir çalışmasının parçası olan yaklaşık 2 metrelik bir timsah mumyası, bir dizi radyografik inceleme için Royal Manchester Çocuk Hastanesi’ne taşındı.
Röntgen ve BT görüntüleri, hayvanın sindirim sisteminde “gastrolit” olarak bilinen küçük taşların olduğunu gösterdi. Timsahlar genellikle yediklerini daha iyi sindirmek ve yüzme dengesini korumak için küçük taşlar yutarlar. Gastrolitlerin bulunması, mumyalayıcıların çürümeyi geciktirmek için iç organları çıkarma işlemi yapmadıklarını gösteriyor.
Sonuçlar, büyük, mumyalanmış timsahların doğadan yakalandığını ve balıkla yemlenmiş oltalar kullanılarak avlandığını gösteriyor. Bu bulgu, MÖ 5. yüzyılda Mısır’ı ziyaret eden Yunan tarihçi Herodot’un timsah avcılığı hakkındaki anlatımını doğruluyor. Herodot, timsahları çekmek için nehir kıyısında domuzların dövüldüğünü ve timsahların Nil’e yerleştirilen yemli oltalarla yakalandığını yazmıştı.Antik Mısır’daki yaşamın birçok yönü hakkında kayıtlar sınırlı olsa da hayvan tapınımı ve mumyalama gibi konularda Mısır’ı ziyaret eden tarihçilerin anlatımları önemli bilgi kaynakları olmaya devam ediyor.
Birmingham Mücevher Okulu’ndaki bazı araştırmacılar ise antik orijinaline benzer bronz bir oltayı yeniden üretip, timsah mumyasının yanında sergileyerek teknolojinin antik geçmişi daha iyi anlamaya nasıl yardımcı olduğunu vurguluyorlar.
Kanser İmmünolojisine Giriş, Neoantijenler, Saklanan Tümörleri Hedef Almak ve Genomik Kanser Aşıları için yorumlar kapalı
Neoantijen nedir? Kanser hastalığına neden olabilecek genetik değişiklikler sonucu yeni tip binlerce farklı protein üretilir ve bunlara neoantijen denir. Parçalanma şeklinde kanser, genlerdeki kusurların bozulması sonucu ortaya çıkar. Bu kadınların çok az bir kısmı “tetikleyicidir”. Bir başka deyişle kanserleşmeye asıl sebep olan bu tip kusurlardır.
Genomik Gardiyan (Koruyucu Melek): p53 Geni ve Proteini Nedir, Genlerimizi Nasıl Korur? için yorumlar kapalı
Genomun gardiyanı olarak da tanımlanan p53 proteini, ilk kez 1979 yılında hayvan deneylerinde incelenen tümörlerde belirlenmiş ve ilerleyen 20 yıl boyunca görevi tam olarak anlaşılamamıştır. Kendisini “gen” olarak tabir etsek de aslında TP53 geni tarafından sentezlenen kısa ömürlü bir proteindir. İlk belirlendiğinde (1979) ağırlığı 53 kDa (kilodalton) olarak bulunduğundan “p53” olarak adlandırılmıştır.
p53, temel görevi hücre bölünmesini düzenlemek olan bir transkripsiyon faktördür. Hücre büyümesinin durdurulması, programlanmış hücre ölümü, hücre farklılaşması ve DNA tamir mekanizmasının başlatılması gibi işlevleri ve birçok organizmada kanserin baskılanmasında rolü olan bir proteindir.
“Genomik gardiyan” olarak anılma sebebi, mutant hücre çoğalmasına karşı genomun korunmasında hücrede oluşmuş bir DNA hasarı tamir olana kadar mitoz bölünmeyi durdurmasıdır.
Genotoksik Stres Yokluğunda Hücre Döngüsü
Sürekli bölünen hücrelerde mitozdan sonra döngü (siklus), G1-S-G2 ve tekrardan mitoz şeklinde tekrarlanarak devam eder. Bu süreçte hücre uyarımı ve büyüme meydana gelmekte veya bölünme sinyali gelmediği sürece istirahat fazı olan G0 evresinde durmaktadır.
G1 fazında hücreler kendi çevrelerini kontrol eder sinyalleri alır ve büyümeyi indükler. Bu fazda DNA sentezi (replikasyon) için hazırlık yapılır, RNA ve protein sentezi yapılır. S fazında, DNA sentezlendikten sonra G2 fazında hücre büyümeye devam ederken yine RNA ve protein sentezi gerçekleşir ve hücre mitoza hazırlanır.
Hücre Döngüsü
Hücre siklusunda bir faz tamamlanmadan sonraki faza geçilirse genetik materyal tam ve doğru kopyalanmadığı için hücrede hasar meydana gelebilir. Hücre siklusunda G1-S ve G2-M geçişinde, ayrıca mitosun safhalarından metafazdan anafaza geçişte kontrol noktaları vardır. Bu kontrol noktalarında hücrenin siklusu devam edip etmeyeceği belirlenir.
Genotoksik Stres Olursa Ne Olur?
Radyasyon veya toksinle karşılaşan hücrelerde -genotoksik stres varlığında- DNA’da meydana gelen hasara göre kontrol noktalarında bir sonraki faza geçiş engellenir. Örneğin, DNA sentezindeki bozulmayı algılayıp p53’ü uyaran protein kinaz ailesinden (PI-3) ATM iyonize radyasyon varlığında, ATR ise UV ışın varlığında p53 genini fosforiller. Tümör baskılayıcı p53 geni işlevini kaybederse hücre büyümesinin kontrolü ortadan kalkar ve DNA tamiri olmadan hücre siklusu kontrolsüz devam eder. Oysa normal hücrelerde DNA hasarı olduğunda, p53 genomik kararlılığı sağlar ve hücre siklusunu G1’de inhibe ederek hücreye tamir için zaman kazandırır. Hasar tamir edilemiyorsa hücre planlı olarak intihar eder (hücrenin planlı olarak kendini öldürmesine “apoptoz” denir.).
p53 geninin birçok kanser türünde mutasyona uğramış olduğu ve normalde hücrede oldukça az miktarda bulunmasına rağmen miktarını arttırdığı bilinmektedir. Kanserin basit bir açıklaması olarak kullandığımız “kontrolsüz hücre çoğalması” tanımının altında yatan sebeptir. Hücre çoğalması p53 geni mutasyonu sonucu kontrol edilemez hale gelir. Dokunun asıl hücrelerine benzeyen yada benzemeyen bir çok hücre oluşur. Çoğalan hücre, köken aldığı dokuya ne kadar benziyorsa kanser o kadar “iyi huylu”dur ve buna “iyi diferansiye” deriz.
Kanser teşhisinde p53 miktarı ve yapısını inceleyip kanserin türü, işleyişi, agresifliği hakkında fikir sahibi olmamız mümkündür. Bu diagnostik avantajı haricinde genomik gardiyanımızın ve onu etkileyen moleküllerin günümüzde yoğun olarak inceleniyor olma sebebi, p53’ün bilim insanlarına kanser tedavilerine “Acaba kanserli dokuda sağlıklı p53 geni aktifleştirilerek hücre çoğalması yeniden kontrol altına alınabilir mi?” sorusu ile gelen alternatif bir bakış açısı sunması olmuştur.
Dinamik metabolomik verilerin analizi, metabolizmadaki altta yatan mekanizmaları anlamamızı geliştirme vaadinde bulunur. Örneğin, bir hastalığın başlangıcından dolayı metabolizmadaki değişiklikleri tespit edebilir. Dinamik veya zamanla çözülmüş metabolomik veriler, bir özne moduna, bir metabolit moduna ve bir zaman moduna göre düzenlenmiş girişlerle üç yönlü bir dizi olarak düzenlenebilir. Bu tür zamanla gelişen çok yönlü veri kümeleri giderek daha fazla toplanırken, bu tür verilerden altta yatan mekanizmaları ve dinamiklerini ortaya çıkarmak zorlu olmaya devam etmektedir. Bu tür veriler için, karmaşıklıklardan biri, çeşitli varyasyon kaynaklarının bir araya gelmesinin varlığıdır: indüklenen varyasyon (deneysel koşullar veya doğuştan gelen hatalar nedeniyle), bireysel varyasyon ve ölçüm hatası. Çok yönlü veri analizi (tensör faktörizasyonları olarak da bilinir), çok yönlü verilerdeki altta yatan kalıpları bulmak için veri madenciliğinde başarıyla kullanılmıştır. Çok yönlü veri analizi yöntemlerinin dinamik metabolomik verilerdeki altta yatan mekanizmaları ortaya çıkarma açısından performansını keşfetmek için, bilinen temel gerçekliğe sahip simüle edilmiş veriler incelenebilir.
Sonuçlar
Artan karmaşıklığa sahip farklı dinamik modellerden, yani basit bir doğrusal sistem, bir maya glikoliz modeli ve bir insan kolesterol modelinden kaynaklanan simüle edilmiş verilere odaklanıyoruz. İndüklenen varyasyonun yanı sıra bireysel varyasyona sahip veriler üretiyoruz. Bu tür dinamik metabolomik verileri analiz etmede çok yönlü veri analizinin avantajlarını ve sınırlamalarını ve farklı varyasyon kaynaklarını çözme kapasitelerini göstermek için sistematik deneyler gerçekleştirilir. Çok yönlü veri analizi yöntemlerinin temel gerçeği bilmenin kolaylaştırdığı yeteneğini anlamak istediğimiz için simülasyonları kullanmayı tercih ediyoruz.
Çözüm
Sayısal deneylerimiz, incelenen dinamik metabolik modellerin giderek karmaşıklaşmasına rağmen, tensör faktörizasyon yöntemleri CANDECOMP/PARAFAC(CP) ve Doğrusal Bağımlılıklara Sahip Paralel Profiller (Paralind)’in varyasyon kaynaklarını çözebileceğini ve böylece altta yatan mekanizmaları ve bunların dinamiklerini ortaya çıkarabileceğini göstermektedir.
Arka plan
Nükleer Manyetik Rezonans (NMR) Spektroskopisi ve Kütle Spektrometrisi (MS) ile gaz kromatografisi (GC) veya sıvı kromatografisi (LC) gibi gelişmiş analitik ölçüm tekniklerinin kullanılabilirliğiyle, biyolojik sistemlerden dinamik veya zamana bağlı (veya uzunlamasına) metabolomik verileri toplamak giderek daha popüler hale geliyor. Bu, bu tür verilerin altta yatan biyolojik süreçleri ve mekanizmaları ortaya çıkarabilme vaadini taşıması nedeniyle daha da popüler hale geliyor. Örnekler, bireylerin sağlık durumlarını araştırmak için meydan okuma testlerinin kullanıldığı metabolizma ve sağlık alanından [ 1 ]; belirli gıda bileşiklerinin metabolik kaderinin incelendiği gıda biliminden [ 2 ]; hastalıkların biyobelirteçlerinin ve hastalık durumlarına erken geçişlerin yakalandığı hastalıkların çalışmasından [ 3 ] vb. verilebilir.
Bahsedilen dinamik metabolomik çalışmalarının temel özellikleri, sınırlı sayıda denekten ölçümlerin alındığı sınırlı sayıda zaman noktası ve farklı varyasyon kaynaklarının üst üste gelmesidir. Farklı varyasyon kaynakları açısından, öncelikle, farklı tedavilerden kaynaklanabilen, örneğin [4]’te ele alınan Qingkailing enjeksiyon grubu veya bir enzimin normalden çok daha düşük aktiviteye sahip olduğu bir hastalıktan kaynaklanabilen, örneğin [ 5 ]’te açıklanan insan mutantları, indüklenmiş varyasyon vardır. İkinci olarak, genellikle oldukça büyük olan bireysel (biyolojik olarak da adlandırılır) varyasyon vardır [ 6 ]. Son olarak, cihaza bağlı olan ve önemli olabilen (kaçınılmaz) ölçüm hatası (teknik hata olarak da adlandırılır) vardır [ 7 ]. Tüm bunlar, bu tür dinamik metabolomik verilerin analizini zorlaştırır.
Bu zorluklar göz önüne alındığında, boyut azaltma yöntemleri, gürültü azaltma (örneğin, ölçüm hatasıyla başa çıkma) ve varyasyonun temel temel kaynaklarını yakalama (dinamik metabolomik verileri analiz etmek için farklı yöntemler hakkında bir inceleme için Smilde ve ark. [ 8 ]’e bakın) için ideal olduklarından umut verici yaklaşımlardır. Boyut azaltma teknikleri, verilerde temelde düşük bir boyutluluk olduğu gerçeğini kullanır ve sözde iki yönlü veriler için bu tür yöntemlerin prototipik örnekleri, örneğin Ana Bileşen Analizi (PCA) ve Ortogonal Kısmi En Küçük Kareler (OPLS), gücünü göstermiştir [ 9 ] ve uzunlamasına metabolomik veri analizi için dinamik olasılıklı PCA’ya uzantılar yapılmıştır [ 10 ]. Veriler özneler , metabolitler ve zaman gibi iki moddan fazla moda sahip olduğunda , verileri iki yönlü bir dizi olarak ele almak yerine çok yönlü bir dizi (daha yüksek dereceli tensör olarak da adlandırılır) oluşturulabilir ve çok yönlü diziler için tensör faktörizasyonları [ 11 , 12 , 13 , 14 ] olarak bilinen boyut azaltma yöntemleri bu tür zamansal verileri analiz etmek için kullanılabilir. Daha önce zamanla değişen metabolomik verileri analiz etmek için kullanılan iki yönlü PCA tabanlı yöntemlerle karşılaştırıldığında, tensör faktörizasyonları tüm modlardaki temel desenleri aynı anda sağlama vaadinde bulunur, örneğin özneler , metabolitler ve zaman modlarındaki desenler. Tensör faktörizasyonları, tartışma izleme [ 15 ], zamansal bağlantı tahmini [ 16 ], veri akışlarının analizi [ 17 ], nörogörüntüleme veri analizi [ 18 , 19 , 20 ] ve elektronik sağlık kayıtlarının analizi [ 21 ] için veri madenciliğinde zamanla değişen verileri analiz etmede başarıyla kullanılmıştır . Ancak, dinamik metabolomik analizde tensör yöntemlerinin kullanımı, yakın zamana kadar bu tür uzunlamasına metabolomik verilerin eksikliği ve yöntemlerin metabolomikteki performansının sınırlı anlaşılması nedeniyle şimdiye kadar sınırlıydı. Bir istisna, sıçanlarda toksikolojik bir hasar üzerinde zaman içindeki tedavilerin etkisini incelemek için ASCA (ANOVA-eşzamanlı bileşen analizi) ile birleştirilmiş CANDECOMP/PARAFAC (CP) [ 22 , 23 ] tensör modelinin kullanılmasıdır [ 24 ].
Şekil 1
Bu makalede, dinamik metabolomik verileri analiz etmede ve altta yatan mekanizmaları ve dinamiklerini ortaya çıkarmada tensör faktörizasyonlarının potansiyelini araştırıyoruz. Bu tür yöntemlerin temel gerçeğini elde etmek ve sınırlamalarını ve avantajlarını incelemek için, yapılandırılmış doğrusal açık bir sistem, maya glikoliz modeli [ 25 ] ve insan kolesterol modeli [ 5 ] dahil olmak üzere artan karmaşıklığa sahip dinamik sistemlerin simülasyonları yoluyla veri üretiyoruz. Hem glikoliz modeli hem de kolesterol modeli in silico modeller. Bu in silico modeller, bir biyolojik sistemin gerçekçi modelleridir ve indüklenen varyasyonun farklı senaryolarını test etmeye olanak tanır. Gerçek verileri daha iyi taklit etmek için, denklemlerdeki kinetik parametreleri rastgele bozarak bu in silico modellere bireysel varyasyonu dahil ediyoruz ve ayrıca belirli parametrelerde bir azalma sağlayarak mutantları, yani indüklenen varyasyonu dahil ediyoruz. Simüle edilmiş verileri, Şekil 1’de gösterildiği gibi , özneler , metabolitler ve zaman modlarına sahip üç yönlü bir dizi olarak düzenliyoruz . Oluşturulan çok yönlü dizi daha sonra CANDECOMP/PARAFAC modeli olarak bilinen en popüler tensör modellerinden biri kullanılarak analiz edilir. CP modeli benzersiz olduğundan (permütasyon ve ölçekleme belirsizliklerine kadar) [ 13 , 27 ], diğer tensör modelleri yerine bu modeli seçtik, örneğin Tucker3 modeli [ 26 ]. Benzersizlik, dinamik metabolomik verileri analiz ederken önemli olan yorumlanabilir desenlere yol açar. Dahası, doğrusal olarak bağımlı faktörlerin varlığında gizli yapıyı CP modelinden daha iyi ortaya çıkarabildiğinden, kısıtlı bir CP modeli, yani Paralind (Doğrusal Bağımlılıklara Sahip Paralel Profiller) modelini [ 28 ] ele alıyoruz.
Yöntemler
Dinamik sistemler ve veri üretimi
Metabolit konsantrasyonlarının dinamikleri, aşağıdaki formdaki diferansiyel denklemlerle modellenebilir:
DXXDT=F(vv):=SSvv,XX(0)=XX0,(1)
Burada vektör metabolit konsantrasyonlarını temsil eder, türev metabolit konsantrasyonlarının zaman içindeki değişimini tanımlar, vektör metabolitler arasındaki reaksiyon akışlarını tanımlar ve matris metabolik ağı tanımlayan stokiyometrik matristir. Matristeki her satır bir metaboliti temsil eder, her sütun bir reaksiyona karşılık gelir ve her giriş, tüketilen metabolit ile negatif bir katsayı elde edilirken üretilen metabolit ile pozitif bir sayı verilecek bir reaksiyondaki bir metabolitin stokiyometrik katsayısını ifade eder. Vektör genellikle kinetik parametrelere sahip metabolitlerin konsantrasyonlarının bir fonksiyonudur.XXDXXDTvvSSSSvv
Doğrusal açık sistem
Akılar konsantrasyonların doğrusal fonksiyonları ise: , o zaman diferansiyel denklem şu şekilde yeniden yazılabilir: . 11 iç metabolit içeren doğrusal bir açık sistem oluşturuyoruz, burada ve boyutunda bir üç köşegen matristir . matrisindeki alt diyagonal elemanlar olarak ve süperdiyagonal elemanlar olması koşuluyla seçilir . Başlangıç değeri olarak ayarlanmıştır. Doğrusal açık sistem hakkında daha fazla ayrıntı Ek dosya 1’de bulunabilir : Bölüm 1. Verileri ürettiğimizde, simülasyonu [0, 0,2] dakikada ele alırız ve için zaman noktalarında çözümü seçeriz . Yol Ek dosya 1’de gösterilmiştir : Şekil S1.F(vv)=AAXX+BBDXXDT=AAXX+BBBB=103×[0,1,0,0,0,0,0,0,0,0,0,0]TAA11×11AA103×[0,2,0,1,0,5,0,3,2,1,3,0,4,1,0,4]T103×[0,3,0,5,2,2,0,3,3,0,5,1,0,2,0,4]TAA(11,11)=−103XX0=[1,1,1,1,1,1,1,1,1,1,1]T(6+5×k)×0,002k=0,1,…,19
Glikoliz modeli
Glikoliz modeli Van Heerden ve arkadaşları tarafından önerilmiştir [ 25 ] ve Denklem ( 1 )’deki doğrusal olmayan terim kinetik denklemleri tanımlayan parametreleri içerir. Bu model açık bir sistemdir ancak ek döngüler nedeniyle doğrusal açık sistemden çok daha karmaşıktır, örneğin metabolit FBP’den enzim PYK’ye ileri beslemeli kontrol döngüsü, ADP-ATP döngüsü ve Ek dosya 1’deki yol grafiğinde gösterilen NADH-NAD döngüsü : Şekil S3; bu model hakkında daha fazla ayrıntı Ek dosya 1’de bulunabilir : Bölüm 2. Verileri ürettiğimizde, [ 25 ]’te dikkate alınan varsayılan başlangıç değerlerini kullanırız. Simülasyonu [0, 0.2]dakikada ele alırızvvDipnot1içinzaman noktalarında çözümü seçin.(6+5×k)×0,002k=0,1,…,19
Kolesterol modeli
Kolesterol modeli van de Pas ve ark. tarafından önerildi [ 5 ] ve bu model için Denklem ( 1 )’deki doğrusal olmayan terim kinetik denklemlerdeki parametreleri içerir. Glikoliz modeline benzer şekilde, bu model de farklı kolesteroller arasında daha fazla döngüye sahip açık bir sistemdir; Ek dosya 1’deki yola bakın : Şekil S8. Model, örneğin ailevi hiperkolesterolemiye (FH), balık gözü hastalığına, Smith-Lemli-Opitz sendromuna (SLOS) ve diğer hastalıklara neden olan mutasyonlar dahil olmak üzere on bilinen mutasyona sahip verilerle doğrulandı [ 5 ]. Her mutasyon için, bazı belirli enzimler normal durumdan çok daha düşük aktivitelere sahiptir. Bu makalede, bu farklı mutant tiplerini indüklenen varyasyonların farklı kaynakları olarak ele alıyoruz. Verileri, [ 5 ] ‘tekiyle aynı başlangıç ayarlarını kullanarak modeli simüle ederek üretiyoruz , yani tüm normal denekler verilen başlangıç metabolit koşullarıyla başlıyor ve mutant denekler normal deneklerin sabit durum koşullarıyla başlıyor. Zaman noktalarını seçme şeklimiz şu şekildedir: [ 5 ]’te kullanılan zaman noktaları için, yani (logspace(0,6,1000)-1)vvDipnot2 , ilk zaman noktasından başlıyoruz ve toplamda 21 zaman noktası elde edene kadar her 24. zaman noktasını seçiyoruzDipnot3 .
Çok yönlü veri analizi
CANDECOMP/PARAFAC (CP) modeli
Bir tensörün poliadik formundan kaynaklanan CP modeli [ 30 ], 1970’te tanıtıldığından beri popüler hale gelmiştir [ 22 , 23 ]. CP çarpanlara ayırma, bir tensörü birinci rütbeli tensörlerin toplamı olarak gösterir (bkz. Şekil 1 ) ve matris Tekil Değer Ayrıştırması’nın (SVD) bir genellemesi olarak görülebilir. Üçüncü dereceden bir tensör in R bileşenli bir CP modeli aşağıdaki gibidir:XX∈RBEN×J×KXX
⟦⟧XX≈X^X^=⟦λλ;AA,BB,CC⟧:=∑R=1RλRAAR∘BBR∘CCR,
Burada birinci rütbe bileşenleri, sırasıyla faktör matrisleri olan ve vektörlerinden oluşmaktadır ve sütunlarıdır; bir skalerdir ve vektör dış çarpımını ifade eder. Bu tanımlamada, vektörü tarafından emildiği varsayılır . İki yönlü veri kümeleri için çoğu boyut indirgeme yönteminin aksine, CP modeli, ek kısıtlamalar getirmeden, hafif koşullar altında permütasyon ve ölçekleme belirsizliklerine kadar benzersizdir [ 13 , 27 ]. Benzersizlik, CP modelinin yorumlanabilir sonuçlar vermesini sağlar ve bu da onu yorumlanabilir veri analizi için çok tercih edilen bir araç haline getirir. Sonuçlar yorumlanırken, deneklerdeki faktör yüklemeleri , metabolitler ve zaman modları her bileşen için birlikte incelenmelidir.AAR,BBRCCRAA∈RBEN×R,BB∈RJ×RCC∈RK×RλR∘AA,BB,CCλλ
CP modeli , aşağıdaki optimizasyon problemini çözerek eksik girdileri olan verileri analiz etmek için de kullanılabilir [ 31 , 32 ]:
⟦⟧dakikaA,B,C‖BB∗(XX−⟦λλ;AA,BB,CC⟧)‖2,
Burada operatör Hadamard ürünüdür, girdileri aşağıdaki gibidir:∗‖.‖BB∈RBEN×J×K
En az bir modda doğrusal olarak bağımlı etkilere sahip varyasyonların temel kaynakları tarafından üretilen desenlere sahip üç yollu veriler için, en uygun CP modeli bu bağımlılıkları göstermelidir ve böyle bir çözüm rütbe eksiktir. Ancak, standart CP modeli verilerdeki gürültü nedeniyle gerçek gizli yapıyı ortaya çıkarmada başarısız olabilir [ 28 ]. Bunun yerine, başlangıçta kısıtlı bir Tucker modeli [ 33 ] olarak tanıtılan Paralind modeli [ 28 ] olarak adlandırılan özel bir CP modeli durumu daha uygundur. Bu model kısmen benzersizdir, yani yalnızca doğrusal olarak bağımsız faktör vektörlerine sahip faktörlerde benzersizliğe sahiptir ancak doğrusal olarak bağımlı faktörlerde benzersiz değildir. Verilerde bulunan örtük doğrusal bağımlılıkları açıkça temsil eder ve böylece gizli yapıyı daha doğru bir şekilde kurtarır. Ayrıca, Paralind modelinde daha az parametre kullanıldığından aşırı uyuma daha az eğilimlidir. İlk modda doğrusal olarak bağımlı faktörlere sahip Paralind modeli aşağıdaki gibi formüle edilebilir:
⟦⟧XX≈X^X^=⟦λλ;AA~,BB,CC⟧=∑R=1RλRA~R∘BBR∘CCR,
Burada ile ve , ve matrisi, doğrusal olarak bağımlı ilişkileri depolayan ‘bağımlılık matrisi’ olarak adlandırılır. Bu modeli Paralind( S , R , R ) ile gösteriyoruz . matrisi şu şekilde verilebilir:AA~=AAHHAA∈RBEN×SHH∈RS×RBB∈RJ×RCC∈RK×RHHHH
HH=[110001].
Sayısal deneyler
Bu bölümde, öncelikle veri setlerini oluşturmak için kullandığımız kurulumları sunacağız ve ardından CP ve Paralind modellerinin altta yatan mekanizmaları ve dinamikleri yakalama açısından performansını göstereceğiz.
Deneysel kurulum ve detaylar
Veri setlerini tanıtmadan önce, öncelikle bireysel ve indüklenen varyasyonları tanımlıyoruz.
Bireysel varyasyon, sabit kinetik parametrelere eklenen rastgele bozulmaları ifade eder. Bireysel varyasyonun seviyesi ( ile gösterilir ) bozulmaların seviyesine bağlıdır. Doğrusal sistem için, bireysel varyasyon, örneğin %1’lik bir seviye içinde süperdiyagonal ve subdiyagonal elemanlara rastgele bozulmalar eklenerek tanıtılır.βDipnotVarsayılan değerlerden 4’ü) ve her sütunun toplamlarının sıfır olarak tutulması, ancakher zaman uygulanırβ=0,01AA(11,11)=−1×103Dipnot5. Glikoliz ve kolesterol modelleri için, kinetik parametrelerin belirli bir seviyesi içinde, örneğin varsayılan değerlerin %2’si içinde () rastgele bozulmalar eklenerek bireysel varyasyon tanıtılır.β=0,02
İndüklenen değişim, belirli bir kinetik parametrede meydana gelen değişimi ifade eder ve indüklenen değişimin seviyesi ( ile gösterilir ) değişimin seviyesine bağlıdır.α
Aşağıdaki iki tür veri kümesini ele alacağız.
Bir kaynaklı indüklenen varyasyona sahip veri kümesi. Bu tür veri kümesi 20 denek içerir:
( Normal denekler) İlk 10 denek, yalnızca seviyesinde bireysel varyasyonla simülasyonların çalıştırılmasıyla elde edilmiştir ;β
( Anormal seviyesinde bireysel varyasyon değerinin %50 oranında azalmasını sağlayan indüklenen varyasyon ile simülasyonlar çalıştırılarak elde edilir (bu denekleri anormal_ A (7,6) denekleri olarak gösteriyoruz ); glikoliz modeli için, VmaxPFK’nin varsayılan değerlerinde %50 oranında azalma vardırβAA(7,6) Dipnot6 (bu konular abnormal_VmaxPFK konuları olarak gösterilir ); kolesterol modeli için, mutant1 (kullanılarak (bu konular abnormal_mutant1 ile gösterilirα=0,62Dipnot(7 konu).
İki kaynaklı indüklenen varyasyonlara sahip veri kümesi. Bu tür veri kümesi 30 denek içerir ve glikoliz ve kolesterol modelleri için üretilir:
( Normal denekler) İlk 10 denek, yukarıda açıklanan normal deneklerle aynı şekilde kullanılarak üretilir .β=0,02
( Anormal denekler) Bir sonraki 10 denek, glikoliz modelinde abnormal_VmaxPFK ( ) ve kolesterol modelinde abnormal_mutant6 ( dir .α=0,50α=0,35β=0,02
( Anormal denekler) Son 10 denek, glikoliz modelinde abnormal_VmaxPYK ( ) ve kolesterol modelinde abnormal_mutant10 ( değeri vardır .α=0,50α=0,95β=0,02
Her veri seti daha sonra özneler , metabolitler ve zaman modları ile üçüncü dereceden bir tensör olarak düzenlenir . Doğrusal açık sistem ve glikoliz modeli tarafından üretilen veri setleri, öznelerin sayısı × 11 metabolit × 20 zaman noktası boyutundadır ve kolesterol modeli tarafından üretilen veri setleri, öznelerin sayısı × 8 metabolit × 21 zaman noktası boyutundadır.
Veri ön işleme
Analizden önce, her üçüncü dereceden tensörü denek moduna göre merkezliyoruz [ 34 ]. Ayrıca, farklı metabolitlerin konsantrasyonları farklı aralıklarda olduğundan, tensör, metabolit modundaki her dilimin karekök ortalama değerine göre metabolit modunda ölçeklenir [ 34 ].
Model seçimi
Farklı modelleri değerlendirirken ve bileşen sayısını belirlerken, özellikle model uyumu, çekirdek tutarlılık tanısı, çapraz doğrulama ve Tucker’ın uyumluluk katsayısı olmak üzere çeşitli tanılamalar kullanırız. Model uyumu (genellikle açıklanan varyans olarak da adlandırılır ) şu şekilde tanımlanır:
Yerleştirmek=100×(1−‖XX−X^X^‖2‖XX‖2),
Burada ve sırasıyla orijinal verileri ve model tarafından veri yaklaşımını ifade eder. %100’lük bir uyum değeri, model tarafından tamamen açıklandığı anlamına gelirken, %100’den küçük bir uyum değeri, artıklarda açıklanamayan bir kısım kaldığı anlamına gelir. Farklı modeller (örneğin, farklı sayıda bileşene sahip modeller) için model uyumunda belirgin bir değişiklik, daha iyi bir model peşinde koşarken dikkate alınması gereken önemli bir kazanımı gösterir.XXX^X^XX
Çekirdek tutarlılık teşhisinin ayrıca bir CP modelindeki bileşen sayısını belirlemek için de yararlı olduğu gösterilmiştir [ 35 ]. Bir CP modelinin çekirdek tutarlılığı, çekirdek dizisinin süperdiyagonallik derecesinin karşılaştırılmasıyla tanımlanırDipnotCP modelinin 8’i ve CP faktörlerini kullanarak Tucker3 modeliyle [ 26 ] verilerin modellenmesiyle elde edilen çekirdek dizisi. %100’e yakın çekirdek tutarlılık değeri uygun bir modeli gösterir ve çok fazla bileşen kullanılırsa düşmesi beklenir.
Son olarak, model seçimi için çapraz doğrulama yoluyla eksik veri tahmin performansını kullanırız. Daha kesin olarak, verilere biraz gürültü ekleriz, yani,
XXgürültü=XX+ηNN‖XX‖‖NN‖,
burada girişleri standart normal dağılımdan rastgele çekilen üçüncü dereceden bir tensördür ve gürültü seviyesidir. Tensör girişlerinin %20’sini rastgele eksik olarak ayarladık, verileri önceden işledik ve eksik girişleri kurtarmak için farklı modeller (yani, CP ve Paralind) kullandık. Farklı rastgele eksik giriş kümelerini kullanarak yöntemlerin performansını değerlendirmek için bu işlemi 20 kez tekrarladık. Daha sonra farklı modellerin performansı, [ 31 ] olarak tanımlanan tensör tamamlama puanı (TCS) kullanılarak değerlendirildiNNη
TÇS=‖(1−BB)∗(XX^−XXgürültü)‖‖(1−BB)∗XXgürültü‖,
Burada Denklem ( 2 ) ile tanımlanır . TCS, bir model için test hatasının bir değerlendirmesi olarak görülebilir ve daha düşük bir değer, modelin verilerdeki temel kalıpları yakalamada daha iyi davrandığını gösterir.BB
CP modelleri iki faktörlü bir dejenerasyondan muzdarip olabilir (dejenerasyon hakkında daha fazla ayrıntı için [ 36 ]’ya bakın). Modelin iki faktörlü bir dejenerasyona sahip olup olmadığını değerlendirmek için Tucker’ın uyumluluk katsayısını (TC ile gösterilir) kullanırız [ 37 ]. i’inci ve j’inci bileşen için TC değeri şu şekilde tanımlanır:
Bu, her moddaki iki bileşenin kosinüs benzerliğinin ( ) çarpımına karşılık gelir. Bu makalede, TC değerini olarak alıyoruz; burada . – 1’e yakın bir TC değeri, geçerli bir model olmayan dejeneratif bir modeli gösterir.CBenJ=AABenTAAJ‖AABen‖‖AAJ‖TC=TCBen0J0|TCBen0J0|=maksimumBen,J|TCBenJ|
Uygulama detayları
CP modelleri , Tensor Toolbox sürüm 3.1’den [ 39 ] cp-opt [ 38 ] ve cp-wopt [ 31 ] (eksik girişleri olan verilere) kullanılarak , sınırlı bellekli BFGS (LBFGS-B) kullanılarak uygulanırDipnot9 optimizasyon algoritması olarak. Zaman modunda negatif olmayan kısıtlamayı uygularız. Paralind modeliDipnot10, Bro ve diğerleri tarafından tanıtılan algoritma kullanılarak yerleştirilir [ 28 ]. Benzersiz modeller elde etmek için, Paralind modelini yerleştirirken metabolitler modunda faktör matrisinin ortogonal ve zaman modunda negatif olmayan olmasını zorunlu kılıyoruz. Yerel minimumlardan kaçınmak için çoklu rastgele başlatmalar kullanılır. Çekirdek tutarlılığının hesaplanması için, N-yollu araç kutusundan corcond fonksiyonunu kullanırız [ 40 ]. Tüm deneyler MATLAB’da (2020a sürümü) gerçekleştirilir.
Paralind(1,2,2) modeli verilerin %98,38’ini açıklıyor ve bu, ekstra kısıtlama nedeniyle CP modelinden biraz daha düşük. Paralind(1,2,2) modelinin özne modu, iki grup arasında net bir ayrım gösteriyor (Şekil 3 b). Metabolitler modundaki ilk bileşenden (Şekil 3 b), M1 , M2 , M3 ve M4 metabolitlerinin büyük mutlak katsayılara sahip olduğunu ve zaman modunda ilk bileşenin bu metabolitlerde gösterilen dinamikleri yakaladığını gözlemliyoruz . Bu bileşen için kalan metabolitlerin katsayıları sıfıra yakındır. İkinci bileşen için, tam tersi şekilde, M7 , M8 , M9 , M10 , M11 ve M5 , M6 metabolitleri büyük katsayılara sahiptir ve bu metabolitlerde gösterilen dinamikler, zaman modunda ikinci bileşen tarafından yakalanır. Ayrıca, metabolitler modundaki her iki bileşenden (Şekil 3 b), Şekil 3 a’da gösterilen bu iki metabolit arasındaki mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan, metabolitler M6 ve M7 arasında bir sıçrama değişimi gözlemliyoruz . Bu değişim, anormal_ A (7,6) deneklerinde nın azalmasından kaynaklanmaktadır ve değişimin model tarafından başarılı bir şekilde yakalanması, Şekil 3 b’de görüldüğü gibi, denek modunda normal (ilk 10 denek) ve anormal_ A (7,6) (son 10 denek) gruplarının başarılı bir şekilde ayrılmasıyla sonuçlanır .AA(7,6)
Şekil 3
Daha büyük bireysel varyasyon dikkate alındığında, rütbe eksikliği ortadan kalkar ve CP modelleri altta yatan örüntüleri daha iyi yakalar, örneğin bkz. Şekil 4 , burada bireysel varyasyon seviyesi ve 2 bileşenli CP modeli için denek modundaki iki bileşenin benzerlik puanı ‘tür . CP modeli verilerin %62,42’sini açıklar. Şekil 4b’de sunulan metabolitler ve zaman modlarındaki ilk bileşenden, M2 hariç tüm metabolitlerin büyük mutlak değerlere sahip katsayılara sahip olduğunu ve zaman modundaki bileşenin tüm metabolitlerde görülen dinamiği bir dereceye kadar yakaladığını gözlemliyoruz. Metabolitler modundaki ikinci bileşenden ( 4b ), M6 , M7 , M8 , M9 , M10 ve M11 metabolitlerinin büyük katsayılara sahip olduğunu ve zaman modunda bu bileşenin bu metabolitlerde gösterilen hızlı düşüşü yakaladığını gözlemliyoruz . M9 , M10 ve M11’deki dinamikler esas olarak ikinci bileşen tarafından yakalanır, ancak metabolitler M5 , M6 , M7 ve M8’deki dinamikler, Şekil 4a’da gösterildiği gibi, zaman modunda iki bileşenin bir karışımıdır . Ayrıca, Şekil 3b’de gösterilen sıçrama değişimine benzer şekilde, metabolitler modunda ikinci bileşende metabolitler M6 ve M7 arasında bir sıçrama değişimi gözlemliyoruz ( 4b ). Bu , Şekil 4a’da gösterilen metabolitler M6 ve M7’deki mavi ve kırmızı çizgilerin değişimiyle tutarlıdır ve anormal_ A (7,6) deneklerinde ‘nın azalmasından kaynaklanmaktadır . Bu nedenle, denek modundaki ikinci bileşenin, Şekil 4c’de gösterildiği gibi, normal ve anormal_ A (7,6) denekleri bir dereceye kadar ayırabilmesi makuldür.β=0,3C12=−0,24AA(7,6)
Şekil 4
Yardımcı deneyler, CP modellerinin davranışının kinetik katsayılara dayandığını göstermektedir. Bazı özel durumlar için, CP modelleri için dejenerasyon gözlemlenmiştir, örneğin, ayarı ve köşegen elemanlarının 1 , 1, 1, 1, 1, 1]\) olarak ayarlandığı bir üç köşegen matris \({\varvec{{A}}} . Böyle bir ayar ve küçük bir bireysel varyasyonla doğrusal açık sistem tarafından üretilen veriler için, örneğin, , CP modeli dejeneratiftir. Ancak, Paralind modeli bu gibi durumlarda da yararlıdır ve altta yatan dinamikleri yakalar; bkz. Ek dosya 1 : Şekil S2.BB=103×[0,5,0,0,0,0,0,0,0,0,0,0]TAA103×[−1,−2,−2,−2,−2,−2,−2,−2,−2,−2,−2]103×[1,1,1,1,1,1,1,1,1,1]β=0,01
Glikoliz modeli
Tek kaynaklı indüklenen varyasyona sahip veri seti
Veriyi, VmaxPFK’nin varsayılan değerinin %50 azalması ve bireysel varyasyonu seviyesindeki indüklenmiş varyasyon olarak ele alıyoruz. Her bir metabolitin zamansal profilleri Şekil 5a’da gösterilmiştir . Tablo 2’ye dayanarak , 2 bileşenli bir CP modeli kullanıyoruz ve doğrusal açık sistemde olduğu gibi, denek modunda rütbe eksikliği gözlemliyoruz . Rütbe eksikliğini hesaba katmak için, bu veri setini analiz etmek için bunun yerine bir Paralind(1,2,2) modeli kullanıyoruz. CP’nin Paralind’e karşı çapraz doğrulama performansı, verilerin %96,05’ini açıklayan Paralind(1,2,2) modelinin bu veri seti için daha iyi bir seçim olduğunu da gösteriyor (bkz. Ek dosya 1 : Şekil S4). İki denek grubu iyi ayrılabilir ve doğrusal sistemle karşılaştırıldığında, Şekil 5b’de gösterilen metabolit modundaki faktör grafiği, ağın karmaşıklığı nedeniyle daha karmaşıktır. β=0,02Tablo 2 Açıklanan varyans (uyum), çekirdek tutarlılık (CC), Tucker’ın uyumluluk katsayısı (TC), ilk iki bileşenin kosinüs benzerlik puanı ( ) denek modunda ve bileşen sayısı ( R ) glikoliz modeli tarafından üretilen verileri analiz etmek için kullanılan CP modelleri için tek kaynaklı indüklenmiş varyasyon ve bireysel varyasyon düzeyindeC12β=0,02
İlk bileşen, Şekil 5a’da gösterilen bu metabolitler arasındaki mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan ve VmaxPFK’nin azalmasından kaynaklanan, biri F6P ve FBP metabolitleri arasında, diğeri PEP ve PYR metabolitleri arasında olmak üzere iki büyük sıçrama değişikliği olduğunu göstermektedir . F6P ve FBP metabolitleri arasındaki değişiklik, Şekil 3’te doğrusal açık sistem için gösterilen değişikliğe benzer şekilde , doğrudan VmaxPFK’nin azalmasına karşılık gelir. PEP ve PYR metabolitleri arasındaki değişiklik, VmaxPFK’nin azalması ve yol grafiğinde gösterilen ileri beslemeli kontrol döngüsünün neden olduğu FBP azalmasından kaynaklanan VmaxPYK enziminin aktivitesinin azalmasına karşılık gelir (Ek dosya 1 : Şekil S3). Metabolitler G6P , F6P , FBP ve PYR ilk bileşende büyük mutlak katsayılara sahiptir ve bu metabolitlerde gösterilen üçüncü zaman noktalarından gelen dinamikler, Şekil 5b’de gösterildiği gibi, zaman modunda ilk bileşen tarafından iyi yakalanmıştır. Metabolitler FBP ve PYR’de gösterilen çıkıntılar, zaman modunda iki bileşenin doğrusal kombinasyonları tarafından yakalanabilir. Ek bir bileşene sahip modeller, örneğin Ek dosya 1’de gösterildiği gibi G6P’deki çıkıntı gibi daha fazla varyansı yakalamak için yararlı olacaktır : Şekil S5. Ancak, dinamik varyasyonların çoğunu yakaladığı ve yorumlanması daha kolay olduğu için Paralind (1,2,2) modelini kullanmayı tercih ediyoruz.
Metabolitler modundaki ikinci bileşen, Şekil 5a’da gösterilen mavi ve kırmızı çizgilerin geçişiyle tutarlı olan metabolitler BPG ve P3G arasında bir sıçrama değişimini gösterir. Bu geçiş, VmaxPYK reaksiyon hızının düşmesiyle oluşan PEP , P2G ve P3G artışından kaynaklanır.Dipnot11 ve anormal_VmaxPFK denekleri için VmaxPFK’nin azalması nedeniyle FBP , TRIO ve BPG’nin azalması. Metabolitler GLCi , BPG ve ACE ikinci bileşende büyük mutlak puanlara sahiptir ve bu metabolitlerin dinamikleri, Şekil 5b’de gösterildiği gibi zaman modunda ikinci bileşen tarafından iyi yakalanmıştır . Metabolitler TRIO , P3G , P2G ve PEP’de gösterilen dinamikler , zaman modunda her iki bileşenin bir karışımıdır.
Daha yüksek düzeyde bireysel varyasyon düşünüldüğünde, özneler modundaki doğrusal bağımlılık zayıflar, örneğin bkz. Ek dosya 1 : Tablo S1, burada bireysel varyasyon düzeyi ve özneler modundaki iki bileşenin kosinüs benzerlik puanı 2 bileşenli bir CP modeli için dir . Bu nedenle Paralind modelleri yerine CP modelleri tercih edilir. Ek dosya 1 : Tablo S1’den, çekirdek tutarlılık değerleri 2 veya 3 bileşenli bir modelin kullanıldığını göstermektedir. 3 bileşenli modeldeki ek faktör yararlı bilgi sağlamadığından 2 bileşenli CP modelini seçtik. 2 bileşenli CP modeli verilerin %54,12’sini açıklıyor. Metabolitler ve zaman modundaki ilk bileşenden (bkz. Şekil 6b ), P3G , P2G ve PEP metabolitlerinin büyük katsayılara sahip olduğunu ve Şekil 6a’da gösterildiği gibi bu metabolitlerin dinamiklerinin yakalandığını gözlemliyoruz. Metabolitler modundaki ikinci bileşenden (bkz. Şekil 6b ), F6P , FBP , TRIO ve BPG metabolitlerinin büyük mutlak katsayılara sahip olduğunu ve bu metabolitlerin çoğunda mavi ve kırmızı çizgilerin ayrılabilir olduğunu gözlemliyoruz . Bu, Şekil 6c’de gösterildiği gibi, normal ve abnormal_VmaxPFK denekleri arasında ikinci bileşen tarafından denek modunda gözlemlenen ayrımla tutarlıdır. Zaman modundaki ikinci bileşen, bu metabolitlerdeki bazı deneklerin gösterdiği dinamikleri yakalar (Şekil 6a ).β=0,36C12=−0,28
Şekil 6
Daha da büyük bireysel varyasyona sahip veriler için, CP modelleri grupları ayıramayabilir. Başarısızlık, (i) bireysel varyasyonun varyansa hakim olmasından kaynaklanır, örneğin Ek dosya 1’e bakın : Şekil S6, burada bireysel varyasyon seviyesi, indüklenen varyasyona eşittir ( ), (ii) bir veya iki deneğin kendine özgü davranış gösterme olasılığı olan sınırlı sayıda denek ( Ek dosya 1’deki BPG profillerine bakın : Şekil S6a) ve böylece ortaklık çıkarmak zorlaşır. Gerçekten de, denek sayısı daha fazla olduğunda (Ek dosya 1’e bakın : Şekil S7), verilerin %66,56’sını açıklayan 3 bileşenli CP modeli, verilerdeki ana değişimi ( VmaxPFK’nin azalması) yakalayabilir ve normal ve anormal_VmaxPFK için bile başarıyla ayırabilir . İdiopatik davranışların daha yaygın hale gelmesi, modellemeyi kolaylaştırmıştır.β=α=0,50β=α=0,50
İki kaynaklı indüklenen varyasyona sahip veri setiseviyesindeki bireysel varyasyonve iki kaynaklı indüklenen varyasyon ile üretilen verileri VmaxPFK için varsayılan değerin %50 azalması ve VmaxPYK için varsayılan değerin %50 azalması. Tablo 3, verilerin %88,68’ini açıklayan 2 bileşenli bir CP modelinin kullanıldığını göstermektedir.β=0,02Tablo 3 Açıklanan varyans (uyum), çekirdek tutarlılık (CC), Tucker’ın uyumluluk katsayısı (TC), ilk iki bileşenin kosinüs benzerlik puanı ( ) denek modunda ve bileşen sayısı ( R ) glikoliz modeli tarafından üretilen verileri analiz etmek için kullanılan CP modelleri için VmaxPFK’de %50 azalma ve VmaxPYK’de %50 azalma olarak iki kaynaklı indüklenmiş varyasyon ve ayrıcaC12β=0,02
Metabolitlerdeki ve zaman modundaki ilk bileşenden (bkz. Şekil 7b ), GLCi , G6P , F6P , FBP , TRIO , BPG , PYR ve ACE metabolitlerinin büyük mutlak katsayılara sahip olduğunu ve Şekil 7a’da gösterilen bu metabolitlerin çoğundaki dinamiklerin yakalandığını gözlemliyoruz. Ayrıca, Şekil 7a’da gösterildiği gibi, mavi çizgiler bu metabolitlerdeki diğer çizgilerden ayrılabilir . Bu, denek modundaki ilk bileşenin (Şekil 7c ) abnormal_VmaxPFK deneklerini diğerlerinden ayırdığı gözlemiyle tutarlıdır. Dahası, Şekil 7a’daki VmaxPFK azalması ve ileri beslemeli kontrol döngüsü nedeniyle mavi çizgilerin diğer çizgilerle yer değiştirmesine uygun olarak, F6P ve FBP metabolitleri ile PEP ve PYR arasında bir sıçrama değişimi gözlemliyoruz . Bu gözlemler, Şekil 5 a, b’de glikoliz modeli için bir kaynaklı indüklenen varyasyon için fark edilenlere benzerdir . Metabolitler ve özneler modundaki ikinci bileşenden (bkz. Şekil 7 b, c), P3G , P2G ve PEP metabolitlerinin büyük puanlara sahip olduğunu ve üç tür öznelerin birbirinden ayrılabileceğini görüyoruz . Bu, Şekil 7 a’da gösterildiği gibi , P3G ve P2G metabolitlerinde farklı renklerde çizgilerin ayrılabilmesi nedeniyle mantıklıdır. Dahası, metabolitler modunda bu bileşende PEP ve PYR metabolitleri arasında bir sıçrama değişimi gözlemliyoruz . Bu, Şekil 7 a’da gösterilen diğer çizgilerle yeşil çizgilerin yer değiştirmesiyle uyumludur ve VmaxPYK’nin azalmasından kaynaklanmaktadır . Zaman modunda, P3G ve P2G metabolitlerinde gösterilen dinamiklerin ikinci bileşen tarafından yakalandığını gözlemliyoruz .
Kolesterol modeli
Tek kaynaklı indüklenmiş varyasyona sahip veri seti İndüklenmiş varyasyona sahip verileri mutant1 ve bireysel varyasyonu. Önceden işlenmiş verilerin zamansal profilleri Şekil 8a’da gösterilmiştir . Ek dosya 1 : Tablo S2, 2 veya 3 bileşenli bir modelin kullanıldığını gösterir ve hem iki hem de üç bileşene sahip CP modelleri için denek modunda rütbe eksikliği gözlemlenirβ=0,02
Şekil 8
Bu nedenle Paralind modelini kullanıyoruz ve yorumdan, 2 bileşenli bir modeli tercih ediyoruz. Dahası, çapraz doğrulama performansı (Ek dosya 1 : Şekil S9), Paralind(1,2,2) modelinin 2 bileşenli CP modelinden daha iyi davrandığını göstermektedir. Paralind(1,2,2) modeli verilerin %89,10’unu açıklamaktadır. Özneler modundaki faktör grafiğinden ( Şekil 8 b), normal ve abnormal_mutant1 özneleri arasında net bir ayrım görüyoruz . Metabolitler ve zaman modundaki ilk bileşenden (bkz. Şekil 8 b), HDL-CE , nonHDL-C ve Per-C metabolitlerinin büyük katsayılara sahip olduğunu, kalan metabolitlerin katsayılarının ise sıfıra yakın olduğunu gözlemliyoruz ; zaman modundaki bileşen, metabolit nonHDL-C’de gösterilen dinamikleri ve ayrıca Şekil 8 a’da gösterildiği gibi HDL-CE ve Per-C metabolitlerindeki dinamiklerin bir karışımını yakalar . Ayrıca, Şekil 8a’daki mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan metabolitler HDL-CE ve nonHDL-C arasında net bir sıçrama değişimi gözlemliyoruz ve bu, mutant1’in neden olduğu abnormal_mutant1 denekleri için metabolitler nonHDL-C’nin yükselmesi ve metabolitler HDL-CE’nin azalmasından kaynaklanmaktadır. Metabolitler ve zaman modundaki ikinci bileşenden (bkz. Şekil 8b ), Liv-FC , Int-FC , Liv-CE ve Int-CE metabolitlerinin büyük katsayılara sahip olduğunu ve zaman modundaki bileşenin , Şekil 8a’da gösterildiği gibi, bu metabolitlerde gösterilen ortak dinamikleri yakaladığını görüyoruz ; ayrıca, Şekil 8a’da gösterilen mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan metabolitler nonHDL-C ve Per-C arasında bir sıçrama değişimi gözlemliyoruz .
Şekil 9
Bu değişim aynı zamanda mutant1’den de kaynaklanmaktadır çünkü nonHDL-C’den Liv-FC ve Per-C’ye olan reaksiyon hızı azalır ve bu da nonHDL-C’nin artmasına ve Per-C’nin azalmasına yol açar .
Yüksek düzeyde bireysel varyasyonlar dikkate alındığında, denekler modundaki rütbe eksikliği ortadan kalkar ve CP modelleri tercih edilir. düzeyinde bireysel varyasyona sahip verileri ele alıyoruz. Ek dosya 1 : Tablo S3’e dayanarak , verilerin %79,15’ini açıklayan 4 bileşenli bir CP modeli kullanıyoruz. Metabolitlerdeki ve zaman modundaki ilk bileşenden (bkz. Şekil 9 a), HDL-CE ve nonHDL-C metabolitlerinin en büyük mutlak katsayılara sahip olduğunu ve Şekil 9 b’de gösterildiği gibi metabolit HDL-CE’deki dinamiklerin yakalandığını gözlemliyoruz . Ek olarak, Şekil 9 c’de denekler modundaki ilk bileşenin normal ve abnormal_mutant1 denekleri ayırdığını görüyoruz . Bu mantıklıdır çünkü mavi ve kırmızı çizgiler Şekil 9 a’da gösterildiği gibi metabolitler HDL-CE ve nonHDL-C’de ayrılabilir. Zaman modundaki ikinci bileşen, metabolitler modunda ikinci bileşende en önemli mutlak puanı alan metabolit HDL-FC’de gösterilen dinamikleri yakalar . Üçüncü bileşen, metabolitler modunda üçüncü bileşende en büyük pozitif puanı alan metabolit Int-CE’de gösterilen dinamikleri yakalar ve dördüncü bileşen , metabolitler modunda dördüncü bileşende en büyük pozitif puanı alan metabolit Per-C’de gösterilen dinamikleri yakalar .β=0,65
Şekil 10
İki kaynaklı indüklenmiş varyasyona sahip veri kümesi Bireysel varyasyonla[ 5 ]’teki mutant6 ve mutant10 olarak iki kaynaklı indüklenmiş varyasyonu. Ek dosya 1 : Tablo S4, 2 bileşenli bir modelin kullanıldığını gösteriyor. İki bileşenli CP modeli verilerin %91,89’unu açıklıyor.Şekil 10c’de gösterilen denek modundan , ilk bileşenin abnormal_mutant10 deneklerini kalan deneklerdenayırdığını , ikinci bileşenin ise normal denekleri abnormal_mutant6 deneklerinden ayırdığını gözlemliyoruz. Bu mantıklıdır çünkü HDL-CE hariç tüm metabolitlerin metabolit modunda ilk bileşen üzerinde büyük bir katsayıları vardır ve Şekil 10a’dan bu metabolitler için önceden işlenmiş verilerdeki mavi çizgilerin ve kırmızı çizgilerin oldukça yakın olduğunu ve yeşil çizgilerden açıkça ayrıldığınıgörebiliriz Metabolit HDL-CE ikinci bileşende en büyük mutlak puana sahipken ve mavi çizgiler,Şekil 10 a’da gösterildiği gibi metabolit HDL-CE için diğer çizgilerden açıkça ayrılabilir. Metabolitlerdeki ve zaman modundaki grafikleri birleştirdiğimizde (Şekil 10 b), modelin iki ana tür dinamiği yakaladığını gözlemliyoruz, yani metabolit HDL-CE’de gösterilen sabit duruma (ikinci bileşen) hızla artanve kalan metabolitlerin çoğunda gösterilen sabit duruma (birinci bileşen) doğru yavaşça artan.β=0,02
Çözüm
Bu makalede, dinamik sistemlerin simülasyonları yoluyla üretilen dinamik metabolomik verilerinin analizi için tensör faktörizasyonlarını inceledik. CP ve Paralind modeli de dahil olmak üzere bu tür yöntemlerin temel fikri, konular arasındaki ortaklığı, yani ortak dinamik davranışları çıkarmaktır. Pratikte karşılaşılan metabolik sistemlerin dinamik davranışı (i) farklı varyasyon kaynaklarının boyutlarına ve (ii) sistemin kendisinin yapısına, yani metabolik ağın topolojisinin yanı sıra kinetik sabitlerin boyutlarına bağlıdır. Artan karmaşıklığa sahip dinamik sistemleri, yani doğrusal açık bir sistem, bir maya glikoliz modeli ve bir insan kolesterol modeli kullanarak, sistemin yapısını ve farklı varyasyon kaynaklarını inceledik ve CP ve Paralind modellerinin farklı ortamlarda altta yatan dinamikleri ne kadar iyi yakaladığını gösterdik. İncelediğimiz yeterli ortaklığın olduğu tüm durumlarda, üç yollu verileri nispeten basit çok yollu modellerle, yani CP ve Paralind modelleriyle modelleyebiliriz. Bu modeller, metabolitler arasındaki ilişkilerdeki değişikliklerin başarılı bir şekilde yakalanmasıyla yansıtılan verilerdeki müdahaleleri tespit etmeyi başarır; bu, metabolitlerin faktör grafiklerindeki sıçrama değişiklikleriyle gösterilir. Metabolik ağ (topoloji ve bağlantı güçleri) ile CP veya Paralind modellerindeki metabolitlerin faktör yüklemeleri arasındaki ilişkinin ayrıntılı bir açıklaması, takip araştırmasının konusudur. Çoğu durumda, altta yatan silico modelden çıkarılan kalıpları da açıklayabilir ve anlayabiliriz. Ancak, dinamik davranıştaki bireysel farklılıklar pratikte, örneğin meydan okuma testlerinde muazzam olabilir [ 41 ]. Bu, örneklenen sınırlı sayıda bireyde, kendine özgü davranışa sahip bazılarının olacağı anlamına gelir. Deneylerimizde, bu kendine özgü davranışın daha çok bir yetersiz örnekleme sorunu olduğunu gösterdik.
CP ve Paralind modeli arasındaki seçim veri özelliklerine bağlıdır ve bu da, yukarıdaki paragrafta tartışılan (i) ve (ii) iki yönüne bağlıdır. Bu makalede, pratikte uygun bir model seçmek için iyi tanılamalar sunuyoruz. Küçük bireysel varyasyona sahip veriler ve dinamik davranış üzerinde benzer etkilere sahip indüklenen varyasyon kaynakları için Paralind modelini kullanıyoruz (CP modelindeki doğrusal bağımlılık faktörleri nedeniyle); büyük bireysel varyasyona sahip veriler veya çeşitli indüklenen varyasyonlara sahip veriler için CP modellerinin iyi çalıştığını gösteriyoruz.
Gecikmeli dinamik sistemler veya indüklenen varyasyondaki önemli farklılıklardan dolayı farklı dinamiklere sahip sistemler veya büyük kendine özgü davranışlar gibi daha karmaşık durumlar için, PARAFAC2 [ 42 ] veya Sınırlı Tucker [ 33 ] gibi daha karmaşık çok yönlü modellere ihtiyaç duyabiliriz. Ayrıca, zamanla evrimleşen metabolitlerle ilgilendiğimiz durumlar için [ 10 ], PARAFAC2’nin metabolitler modunda evrimleşen faktör matrislerini yakalayarak bunları ortaya çıkarması beklenir . Ayrıca, bireyler arasındaki rastgele varyasyonu hesaba katan karışık etkili üç yönlü modelleri dikkate almaya değer olabilir.
Bu simülasyon çalışması gerçek bir dinamik metabolomik veri setinin analizinden ilham almıştır. Gerçek verilerde, altta yatan dinamik ağ bilinmemektedir ve veri seti boyutu daha büyüktür, örneğin, metabolit ve denek sayısı yüzlerce mertebesindedir. CP modellerinin hala varyasyonların ana modellerini ve karşılık gelen zamansal profilleri ortaya çıkarması beklenmektedir, bunu gerçek bir metabolomik meydan okuma testi veri setindeki bulgularımızla göstermeyi planlıyoruz. Gelecekte bu tür büyük ölçekli dinamik metabolomik veri setleri mevcut olacaksa, yöntemler daha büyük veri setlerine [ 43 , 44 ] (her modda binlerce veya daha fazla değişkenle) de ölçeklenebilir.
Glikoliz modelindeki reaksiyonlar çok hızlıdır ve metabolitlerin konsantrasyonları sabit duruma hızla ulaşır. Bu nedenle, dinamik değişimin ortaya çıktığı kısa bir zaman aralığına odaklanıyoruz. Ancak, böyle bir zaman ölçeğinde gerçek metabolomik verileri edinmek mümkündür; hücre içi metabolitleri çıkarırken örnekleme süresinin örnek başına 220 ms olabileceği [ 29 ]’a bakın.
MATLAB gösteriminde: tspan=logspace(0,6,1000)-1 vektörünü ele alalım; seçilen zaman noktaları tspan(1:24:500)’dir.
Küçük biyolojik varyasyonlara sahip sistemleri taklit etmek için küçük sayılar seçiyoruz, ancak bu sayı, makalede daha sonra tartışılacağı üzere büyük de olabilir.
Bu kısıtlamalar doğrusal açık sistemlerde kütle korunum yasasının sağlanması için veri üretiminde her zaman kullanılır.
Burada, VmaxPFK dışındaki enzimler de düşünülebilir. Yolun orta kısmında konumlanmış bir enzimle başlamak istediğimiz için VmaxPFK’yi seçiyoruz.
İndüklenen varyasyon diğer enzim reaksiyon hızlarındaki azalmayla, örneğin VmaxPYK / mutant6 / mutant10’un %50 azalmasıyla tanımlanıyorsa , anormal denekler sırasıyla abnormal_VmaxPYK / abnormal_mutant6 / abnormal_mutant10 olarak gösterilir .
Bir CP modelinin çekirdek dizisi, CP modelini Tucker3 modelinin özel bir durumu olarak ifade ederek elde edilen çekirdek tensörüdür. CP modelinin çekirdek dizisi, , yani, rütbe bir bileşenlerinin ağırlıkları, süperdiyagonalde ve diğer tüm girişler sıfır olan bir süperdiyagonal tensördür.λ
VmaxPYK reaksiyon hızının azalması, VmaxPFK’nin azalmasından ve bunun sonucunda FBP’nin azalmasından ve ileri beslemeli kontrol döngüsünün etkisinden kaynaklanmaktadır.
Referanslar
Pellis L, van Erk MJ, van Ommen B, Bakker GC, Hendriks HF, Cnubben NH, Kleemann R, van Someren EP, Bobeldijk I, Rubingh CM, ve diğerleri. Yemek sonrası bir meydan okumadan sonra plazma metabolomikleri ve proteomik profillemesi, insan metabolik durumu üzerinde ince diyet etkilerini ortaya koymaktadır. Metabolomik. 2012;8(2):347–59.MaddeCASGoogle Akademik
van Duynhoven J, Vaughan EE, Jacobs DM, Kemperman RA, van Velzen EJ, Gross G, Roger LC, Possemiers S, Smilde AK, Doré J, ve diğerleri. İnsan süperorganizmasındaki polifenollerin metabolik kaderi. Proc Natl Acad Sci. 2011;108(Ek 1):4531–8.MaddeGoogle Akademik
Price ND, Magis AT, Earls JC, Glusman G, Levy R, Lausted C, McDonald DT, Kusebauch U, Moss CL, Zhou Y, ve diğerleri. Kişisel, yoğun, dinamik veri bulutları kullanan 108 bireyden oluşan bir sağlık çalışması. Nat Biotechnol. 2017;35(8):747.MaddeCASGoogle Akademik
Lin Z, Zhang Q, Dai S, Gao X. Grup ve nükleer norm düzenlenmiş çok değişkenli regresyon yoluyla uzunlamasına hedefsiz metabolomik verilerde zamansal kalıpların keşfi. Metabolitler. 2020;10(1):33.MaddeCASGoogle Akademik
van de Pas NC, Woutersen RA, van Ommen B, Rietjens IM, de Graaf AA. İnsanlarda plazma kolesterol konsantrasyonlarını tahmin eden fizyolojik temelli bir siliko kinetik modeli. J Dudak Res. 2012;53(12):2734–46.MaddeGoogle Akademik
Adamko D, Rowe BH, Marrie T, Sykes BD, ve diğerleri. Normal insan idrarındaki metabolitlerin değişimi. Metabolomik. 2007;3(4):439–51.MaddeGoogle Akademik
Van Batenburg MF, Coulier L, van Eeuwijk F, Smilde AK, Westerhuis JA. Kapsamlı fonksiyonel genomik veriler için yeni değerler: metabolomik durum. Anal Kimya 2011;83(9):3267–74.MaddeGoogle Akademik
Smilde A, Westerhuis J, Hoefsloot H, Bijlsma S, Rubingh C, Vis D, Jellema R, Pijl H, Roelfsema F, Van Der Greef J. Dinamik metabolomik veri analizi: öğretici bir inceleme. Metabolomik. 2010;6(1):3–17.MaddeCASGoogle Akademik
Yamamoto H, Yamaji H, Abe Y, Harada K, Waluyo D, Fukusaki E, Kondo A, Ohno H, Fukuda H. Gizli değişkenlere farklı cezalar veren pca, pls, opls ve rfda kullanılarak metabolom verileri için boyutsallık indirgeme. Chemom Intell Lab Syst. 2009;98(2):136–42.MaddeCASGoogle Akademik
Nyamundanda G, Gormley IC, Brennan L. Uzunlamasına metabolomik verilerin analizi için dinamik olasılıklı ana bileşenler modeli. JR Stat Soc Ser C Appl Stat. 2014;63(5):763–82.MaddeGoogle Akademik
Smilde A, Bro R, Geladi P. Çok yönlü analiz: kimyasal bilimlerdeki uygulamalar. Chichester: Wiley; 2004.KitapGoogle Akademik
Acar E, Yener B. Gözetimsiz çok yönlü veri analizi: bir literatür taraması. IEEE Trans Knowl Data Eng. 2009;21(1):6–20.MaddeGoogle Akademik
Kolda TG, Bader BW. Tensör ayrıştırmaları ve uygulamaları. SIAM Rev. 2009;51(3):455–500.MaddeGoogle Akademik
Papalexakis EE, Faloutsos C, Sidiropoulos ND. Veri madenciliği ve veri füzyonu için tensörler: modeller, uygulamalar ve ölçeklenebilir algoritmalar. ACM Trans Intell Syst Technol. 2016;8(2):16.Google Akademik
Bader BW, Berry MW, Browne M. PARAFAC kullanılarak Enron e-postasında tartışma takibi. Londra: Springer; 2008. s. 147–63.
Dunlavy DM, Kolda TG, Acar E. Matris ve tensör faktörizasyonlarını kullanarak zamansal bağlantı tahmini. ACM TKDD. 2011;5(2):10.Google Akademik
Sun J, Papadimitriou S, Philip SY. Yüksek boyutlu ve çok yönlü akışlarda pencere tabanlı tensör analizi. Veri madenciliği üzerine altıncı uluslararası konferansta (ICDM’06). IEEE; 2006. s. 1076–80.
Acar E, Aykut-Bingol C, Bingol H, Bro R, Yener B. Epilepsi tensörlerinin çok yönlü analizi. Biyoenformatik. 2007;23(13):10–8.MaddeGoogle Akademik
Davidson I, Gilpin S, Carmichael O, Walker P. fMRI verilerinin kısıtlanmış tensör analizi yoluyla ağ keşfi. KDD’13: 19. ACM SIGKDD bilgi keşfi ve veri madenciliği uluslararası konferansının bildirileri. ACM; 2013. s. 194–202.
Roald M, Bhinge S, Jia C, Calhoun V, Adali T, Acar E. Parafac2 modelini kullanarak ağ evriminin izlenmesi. In: ICASSP’20: 45. IEEE akustik, konuşma ve sinyal işleme uluslararası konferansının bildirileri; 2020.
Yin K, Afshar A, Ho JC, Cheung WK, Zhang C, Sun J. Logpar: eksik değerlere sahip zamansal ikili veriler için lojistik parafac2 çarpanlarına ayırma. In: KDD’20: 26. ACM SIGKDD bilgi keşfi ve veri madenciliği uluslararası konferansının bildirileri; 2020.
Harshman RA. PARAFAC prosedürünün temelleri: “açıklayıcı” çok modlu faktör analizi için modeller ve koşullar. UCLA Work Pap Phonet. 1970;16:1–84.
Carroll JD, Chang JJ. “Eckart-young” ayrıştırmasının n-yollu genellemesi yoluyla çok boyutlu ölçeklemedeki bireysel farklılıkların analizi. Psychometrika. 1970;35:283–319.
Jansen JJ, Bro R, Hoefsloot HC, van den Berg FW, Westerhuis JA, Smilde AK. Parafasca: Asca, metabolik parmak izi verilerinin analizi için parafac ile birleştirildi. J Kemom. 2008;22(2):114–21.MaddeCASGoogle Akademik
van Heerden JH, Wortel MT, Bruggeman FJ, Heijnen JJ, Bollen YJ, Planqué R, Hulshof J, O’Toole TG, Wahl SA, Teusink B. Geçişte kayıp: glikolizin başlatılması, büyümeyen hücrelerin alt popülasyonlarını verir. Bilim. 2014;343:6174.
Tucker LR. Üç modlu faktör analizi üzerine bazı matematiksel notlar. Psychometrika. 1966;31(3):279–311.MaddeCASGoogle Akademik
Kruskal JB. Üç yollu diziler: üç doğrusal ayrıştırmaların sıralaması ve benzersizliği, aritmetik karmaşıklık ve istatistiklere uygulama. Doğrusal Cebir Uygulaması 1977;18(2):95–138.MaddeGoogle Akademik
Bro R, Harshman RA, Sidiropoulos ND, Lundy ME. Doğrusal olarak bağımlı yüklemelerle çok yönlü verilerin modellenmesi. J Chemom. 2009;23(7–8):324–40.MaddeCASGoogle Akademik
Schaefer U, Boos W, Takors R, Weuster-Botz D. Hücre içi metabolit dinamiklerini izlemek için otomatik örnekleme cihazı. Anal Biochem. 1999;270(1):88–96.MaddeCASGoogle Akademik
Hitchcock FL. Bir tensörün veya poliadiğin ürünlerin toplamı olarak ifadesi. J Math Phys. 1927;6(1–4):164–89.MaddeGoogle Akademik
Acar E, Dunlavy DM, Kolda TG, Mørup M. Eksik veriler için ölçeklenebilir tensör faktörizasyonları. Chemom Intell Lab Syst. 2011;106(1):41–56.MaddeCASGoogle Akademik
Tomasi G, Bro R. Parafac ve eksik değerler. Chemom Intell Lab Syst. 2005;75(2):163–80.MaddeCASGoogle Akademik
Kiers HA, Smilde AK. İkinci dereceden enstrümantal verilerle parametre tahmini için bir araç olarak kısıtlanmış üç modlu faktör analizi. J Chemom. 1998;12(2):125–47.MaddeCASGoogle Akademik
Bro R, Smilde AK. Bileşen analizinde merkezleme ve ölçekleme. J Chemom. 2003;17(1):16–33.MaddeCASGoogle Akademik
Bro R, Kiers HA. Parafak modellerindeki bileşen sayısını belirlemek için yeni ve etkili bir yöntem. J Chemom. 2003;17(5):274–86.MaddeCASGoogle Akademik
Stegeman A. Candecomp/parafac ve indscal’daki dejenerasyon, iki değerli tipik bir sıralamaya sahip birkaç üç dilimli dizi için açıklandı. Psychometrika. 2007;72(4):601–19.MaddeGoogle Akademik
Bro R. Parafac öğretici ve uygulamaları. Chemom Intell Lab Syst. 1997;38(2):149–72.MaddeCASGoogle Akademik
Acar E, Dunlavy DM, Kolda TG. Kanonik tensör ayrıştırmalarına uyum sağlamak için ölçeklenebilir bir optimizasyon yaklaşımı. J Chemom. 2011;25(2):67–86.MaddeCASGoogle Akademik
Bader BW, Kolda TG, ve diğerleri. Genel yazılım, son sürüm. MATLAB için Tensor Toolbox, Sürüm 3.1.
Andersson CA, Bro R. Matlab için n-yollu araç kutusu. Chemom Intell Lab Syst. 2000;52(1):1–4.MaddeCASGoogle Akademik
Wopereis S, Stroeve JH, Stafleu A, Bakker GC, Burggraaf J, van Erk MJ, Pellis L, Boessen R, Kardinaal AA, van Ommen B. Sağlıklı ve tip 2 diyabetik deneklerde standartlaştırılmış karışık yemek tolerans testinin çok parametreli karşılaştırması : phenflex mücadelesi. Genler Nutr. 2017;12(1):1–14.MaddeGoogle Akademik
Harshman RA. PARAFAC2: matematiksel ve teknik notlar. UCLA Çalışma Pap Phonet. 1972;22:30–47.Google Akademik
Bro R, Andersson CA. Çok yönlü algoritmaların hızının artırılması: Bölüm II: sıkıştırma. Chemom Intell Lab Syst. 1998;42(1–2):105–13.MaddeCASGoogle Akademik
Beutel A, Talukdar PP, Kumar A, Faloutsos C, Papalexakis EE, Xing EP. Flexifact: Hadoop’ta bağlı tensörlerin ölçeklenebilir esnek faktörizasyonu. In: 2014 SIAM uluslararası veri madenciliği konferansı bildirileri; 2014.
Jendoubi T, Ebbels TMD. Zaman seyri metabolik verilerinin ve biyobelirteç keşfinin bütünleştirici analizi. BMC Bioinform. 2020;21:11.MaddeGoogle Akademik
Acar E, Bro R, Smilde AK. Eşleştirilmiş matris ve tensör faktörizasyonlarını kullanarak metabolomikte veri birleştirme. IEEE Bildirileri. 2015;103:1602–20.MaddeCASGoogle Akademik
Teşekkürler
Amsterdam Üniversitesi Swammerdam Yaşam Bilimleri Enstitüsü’nden Dr. Meike T. Wortel’e, maya glikoliz modeli hakkındaki birçok yararlı iletişimi ve içgörüsü için teşekkür etmek istiyoruz. Ayrıca, Danimarka Pediatrik Astım Merkezi’ndeki (COPSAC) Dr. Morten Arendt Rasmussen ve işbirlikçilerimize de yararlı tartışmalar için minnettarız. Ayrıca, makalemizi iyileştirmemize yardımcı olan değerli yorumları için değerlendiricilere de minnettarız.
Finansman
Bu makalede sunulan çalışma Novo Nordisk Vakfı Hibe NNF19OC0057934 ve Norveç Araştırma Konseyi proje #300489 tarafından desteklenmiştir.
Geleceğin Ayak Sesleri: Nanoteknoloji için yorumlar kapalı
Yuri N. Artsutanov bunu düşledi. Hem de 1960’lı yıllarda. Peki böyle bir şey gerçekten mümkün mü? Şimdilik teoride olsa da konuyla ilgili çalışmalar çoktan başladı. Bu hayali gerçekleştirmek için en önemli aşama, belki de insanlığın bildiği en sağlam malzemeyi üretebilmekte. Bunun yolu da nanoteknolojiden geçiyor. Bilimle ilgisi olsun olmasın hemen herkes, nanoteknoloji sözcüğünü bir yerlerden duymuştur. Her geçen gün hayatımıza daha fazla giren bu kavramın ne olduğunu hiç merak ettiniz mi? Gözlerinizin göremeyeceği kadar küçük bir dünyanın içinde, kısa bir yolculuğa çıkalım.
Nano sözcüğü etimolojik olarak Yunanca kökenli ve aslında “cüce” anlamına geliyor. Bugün bu sözcüğü, metrenin milyarda biri ölçülerine denk gelen fiziksel boyutları tanımlamakta kullanıyoruz. “Boyutları 1 ile 100 nanometre arasında değişen maddeler üzerinde yapılan çalışmalar da nanobilim ve nanoteknoloji dediğimiz alanları oluşturuyor.” Başka bir deyişle, maddenin atomik ve moleküler düzeyde işlenmesi… Evet, söz konusu nanoboyut olduğunda, gerçekten çok çok küçük şeylerden bahsetmeye başlıyoruz. Aşağıdaki tablo nanoboyutun daha iyi anlaşılması için yararlı olacaktır. Zira saç telinin yarıçapının bile devasa sayılabileceği bir oran görüyoruz:Peki ama maddeyi atom ve molekül boyutunda kontrol edebilme fikri nereden çıktı?
1959 yılında ünlü fizikçi Richard Feynman, “There’s Plenty Of Room at the Bottom” adlı bir konuşma yaptı. Bu meşhur konuşmada Feynman, gelecekte çok küçük boyutlarda maddelerin kontrol edilip bunların üzerinde çalışmalar yapılabileceğini öngörüyordu. Bu ilham verici tahminler, yıllar sonra gerçeğe dönüşmeye başladı. Aslında nanoteknoloji sözcüğü 1974’e kadar hiç kullanılmamıştır. Japon bilim insanı Norio Taniguchi’nin bu tarihte bir konferansta nanoteknoloji teriminden bahsettiğini görüyoruz. Ancak bu terimin popülerleşmesi başka bir isim sayesinde oldu: K. Eric Drexler, 1976 yılında yayımlanan Engines of Creation: The Coming Era Of Nanotechnology adlı kitabında bu yeni dünyaya adeta hoş geldin diyordu. Bugün, canlı vücuduna enjekte edilebilecek nanorobotlar üzerine araştırmalar yapılıyorsa; tüm bu gelişmelerde, yıllar önce yaşamış öncülerin payları yadsınamaz. İnsanlık, onların gösterdiği yol boyunca, uzay araştırmalarından gıda endüstrisine, savaş sanayinden sağlık sektörüne, pek çok alanda yeni keşiflere imza attı. Örneklerle devam edecek olursak, Yıllar önce geçirdiği bir trafik kazası sonucunda felç geçirerek uzuvlarının kontrolünü yitiren Ian Burkhart adlı bir hasta; beynine yerleştirilen nanoteknoloji ürünü küçük bir mikroçip sayesinde artık kollarını kullanabiliyor. Kendi kendini temizleyebilen yüzeyleri duymuşsunuzdur. Ya da yıkamaya gerek duyulmayan giysileri. Fotokatalizle parçalanan kirlerin ardından el sallıyorsunuz. Sonra da nanoteknoloji ürünü yarı iletken malzemelere teşekkür ediyorsunuz. Tabii yeterince kibarsanız… Nanoteknoloji her yıl daha parlak fikirlerle gelişiyor. İlerleyen yıllarda bizleri çok büyük sürprizlerin beklediğinden şüphemiz yok. Yazının başında değindiğimiz uzay asansörü fikri, uzay yolculukları ve yörüngeye nakliye maliyetleri açısından büyük önem taşıyor. Bunun gibi olağanüstü projeler belki nanorobotlar sayesinde gelecekte sıradanlaşacak. Vücudunuzda, hücreleriniz gibi uyum içinde çalışan küçük robotların bulunduğunu hayal edin. Hastalıklarınızı erkenden algılayıp tedavi edebilecek minik doktorlar. Besin ihtiyacınızı minimalize etmek için çabalayacak mikro aşçılar. İletişim gereksiniminizi cihaz ve aksesuarlardan arındıracak küçük asistanlar…
Bilimin Doğasını Anlamak: Evrim Eğitiminde Bir Önkoşul için yorumlar kapalı
Bilimde ilerleme doğal olayların nedenleri hakkında daha iyi açıklamaların geliştirilmesi ile olur. Yani bilimde ortaya konulan bir açıklamanın tam ve kesin olduğundan emin olmak zordur. Ancak pek çok bilimsel açıklama hakkında o kadar çok bilgi toplanmış ve bu açıklamalar o kadar çok testten geçmiştir ki bugün bu açıklamaların güvenilirliği çok yüksektir.
Programlanmış ölüm-1 (PD-1), T hücresi yanıtını baskılayan bir bağışıklık kontrol noktası molekülüdür. PD-1’in PD-L1/PD-L2’ye bağlanması sitokin üretimine neden olur ve T hücresi çoğalması azalır. PD-L1 ve PD-L2 ifade eden tümörler sitotoksik T hücrelerinden kaçar ve tümör ilerlemesine maruz kalır. Bu nedenle, immünoterapi kanser tedavisinde yeni bir seçenek haline gelmiştir.
Amaçlar:
Bu çalışmada kolorektal karsinomda (KRK) PD-L1 ve PD-L2 ekspresyonunu inceledik ve klinikopatolojik parametreler ile CD8+ T hücreleri arasındaki ilişkiyi değerlendirdik.
Yöntem ve Materyal:
Kolorektal kanser tanısı almış 124 hastaya ait formalin fiksasyonu yapılmış parafine gömülmüş tümör hücreleri ve immün hücrelerde PD-L1, PD-L2 boyama yöntemi kullanılarak tümör infiltre eden lenfositlerde ve çevresindeki tümör lenfositlerinde CD8 ekspresyonu değerlendirildi.
Kullanılan İstatistiksel Analiz:
Çapraz tabloların analizinde Pearson Ki-Kare, Fisher Tam Ki-Kare ve Pearson Tam Ki-Kare analizleri kullanıldı. Sağkalım dağılımları Kaplan-Meier yöntemini tahmin etti ve log-rank istatistikleri kullanılarak değerlendirildi.
Sonuçlar:
Çalışmamızda PD-L1 ekspresyonu ile kadın cinsiyeti ve medüller morfolojiye sahip tümörler arasında anlamlı bir korelasyon bulundu. Medüller morfolojiye sahip tümörlerde PD-L2 ekspresyonu gözlenmedi ve PD-L2 ile medüller bileşen arasında istatistiksel olarak ters bir ilişki gözlendi. PD-L1 pozitif tümör infiltre eden lenfositlerin nükssüz sağ kalım için önemli bir öngörücü olduğu belirlendi.
Kolorektal karsinom (CRC), dünya çapında erkeklerde ve kadınlarda en sık görülen üçüncü kanserdir.[ 1 , 2 ] CRC’de cerrahi, kemoterapi, radyoterapi ve immünoterapi veya kombinasyon tedavileri tercih edilir. Ancak, immünosupresyonla oluşan tümör mikroçevresi nedeniyle, bu tedaviler farklı hastalarda farklı sonuçlara sahiptir. Bu durum, CRC’li hastalarda kişiselleştirilmiş tedavi yöntemlerinin geliştirilmesini gerektirir.[ 2 , 3 ]
Programlanmış ölüm reseptörü-1 (PD-1), CD28/CTLA-4 ko-stimülatör reseptör ailesinin bir üyesidir ve T hücresi düzenleyicilerinden biridir. T hücresi, B hücresi ve doğal öldürücü hücre yüzeylerinde ifade edilir. Programlanmış ölüm ligandı-1 (PD-L1, B7-H1 ve CD274) ve programlanmış ölüm ligandı-2 (PD-L2, B7-DC), tümör hücreleri, T ve B hücreleri, makrofajlar ve dendritik hücreler gibi birçok hücrede ifade edilen bu reseptörün ligandlarıdır. PD-1 ve PD-L1/L2’nin bağlanması, inhibe edici bir sinyale yol açar. Sonuç olarak, sitokin üretimi ve T hücresi çoğalması azalır.[ 4 , 5 ] Melanom, küçük hücreli olmayan akciğer karsinomu ve böbrek hücreli karsinomlarda anti-PD-1 ve anti-PD-L1 ile olumlu sonuçlar elde edilir. Gastrointestinal maligniteli hastalarda PD-L1 ekspresyonu ve prognozla ilişkisi belirsizdir.[ 6 , 7 , 8 ] Mevcut çalışma, CRC’de PD-L1 ve PD-L2 ekspresyonunu incelemeyi ve klinikopatolojik parametreler ile CD8+ T hücreleri arasındaki ilişkiyi değerlendirmeyi amaçlamaktadır.
KONULAR VE YÖNTEMLER
Hastalar ve tümör özellikleri
Çalışmaya 2010-2013 yılları arasında Tıp Fakültesi Patoloji Anabilim Dalı’nda Kolorektal Kanser tanısı almış 124 hasta dahil edildi. Tüm olgular rezeksiyon materyalleri olan hastalardan seçildi. Endoskopik biyopsi materyalleri, nüks tümörler ve neoadjuvan tedavi alan hastalar çalışmaya dahil edilmedi. Demografik özellikler, tümör lokalizasyonu ve tümör boyutu kaydedildi. Olguların patolojik preparatları arşivden elde edilerek histolojik tip, diferansiyasyon, nekroz, küçük damar invazyonu, büyük damar invazyonu, perinöral invazyon varlığı, invazyon derinliği ve metastatik lenf nodu sayısı açısından tekrar değerlendirildi. Değerlendirme klinik dosyalardan yapılarak takip aralıkları ve nüks süresi elde edildi.
İmmünohistokimyasal boyama ve değerlendirme
Bu çalışmada tümörü temsil eden parafin bloklar seçilerek tümör etrafındaki inflamatuar hücreler değerlendirildi ve en yoğun olanlar seçildi.
Bloklardan dört mikron kalınlığında kesitler hazırlandı. PD-L1 (seyreltme 1:100, tavşan monoklonal, ab205921; Abcam, Cambridge, MA), PD-L2 (seyreltme 1:300, tavşan monoklonal, klon D7U8C, Cell Signaling Technology, Danvers, MA) ve CD8 (SP16 klonu, Neo belirteci) (uygulanmaya hazır formda) seyreltilmeden dokulara uygulandı. DAKO Omnis otomatize immünohistokimyasal boyama cihazı (DAKO Omnis, Danimarka) kullanılarak doku kesitleri hazırlandı. Kesitler damıtılmış su, iki kez %96’lık alkol, bir kez %99’luk alkol ve üç kez ksilen ile yıkandı. Boyanmış preparasyonlar ksilen bazlı slayt kapatıcı ile kapatılarak değerlendirmeye hazır hale getirildi. PD-L1 ile membranöz boyama pozitif olarak değerlendirildi. Tümör hücrelerinin ve bağışıklık hücrelerinin boyanması değerlendirildi. Her iki grup için de %5 ve üzeri PD-L1 ekspresyonu pozitif, %5’in altı ise negatif olarak kabul edildi.[ 6 ] Tümör hücrelerinin sitoplazmik ve/veya membranöz boyanması PD-L2 ile immünohistokimyasal analizde pozitif olarak kabul edildi. Tümör hücrelerinin boyanma yoğunluğu hafif, orta ve şiddetli olmak üzere üç gruba ayrıldı. Orta ve şiddetli yoğunluklarda %10 ve üzeri pozitif, %10’dan azı ise negatif olarak kabul edildi. Hafif yoğunlukta %50 ve üzeri boyanma pozitif, %50’nin altındaki boyanma ise negatif olarak kabul edildi.[ 9 , 10 ]
CD8 boyama, tümör infiltre eden lenfositler (TIL) ve çevreleyen tümör lenfositleri olarak iki farklı kategoride değerlendirildi. Tümör içerisinde ×200 amplifikasyon alanında 50 ve üzeri CD8 pozitif lenfosit varlığı yüksek, 50’den az CD8 pozitif lenfosit varlığı düşük olarak kabul edildi. Tümör içerisinde ×200 amplifikasyon alanında 200 CD8 pozitif lenfosit varlığı yüksek, 200’ün altında lenfosit varlığı düşük olarak kabul edildi.[ 11 ] İki patolog, klinikopatolojik verilerden habersiz olarak immünohistokimyasal boyamayı bağımsız olarak değerlendirdi.
İstatistiksel analiz
Sürekli veriler ortalama ± standart sapma ve medyan değer (%25-75) olarak verildi. Nitel veriler ise yüzde (%) olarak verildi. Verilerin normal dağılıma uygunluğu Shapiro-Wilk testi ile araştırıldı. Normal dağılım göstermeyen iki grup karşılaştırıldığında Mann-Whitney U testi kullanıldı. Çapraz tabloların analizinde Pearson Ki-Kare, Fisher Kesin Ki-Kare ve Pearson Kesin Ki-Kare analizleri kullanıldı. Sağkalım dağılımları Kaplan-Meier yöntemini öngördü ve log-rank istatistikleri kullanılarak değerlendirildi. Olaya kadar geçen süre kolon rezeksiyon tarihinden olay tarihine kadar geçen süre olarak hesaplandı. Karıştırıcı faktörleri düzeltmek için geriye doğru koşullu yöntemi kullanan Cox regresyon modeli kullanıldı. Analizde IBM SPSS Statistics 21.0 programı kullanıldı. İstatistiksel anlamlılık için P <0,05 kriter olarak kabul edildi. Çalışmamız Eskişehir Osmangazi Üniversitesi Tıp Fakültesi İlaç Dışı Klinik Araştırmalar Etik Kurulu’ndan onaylandı (Tarih:14/11/2016, No:02).
SONUÇLAR
Hastanın özellikleri
Çalışma kohortunun ( n = 124) ortalama yaşı 64 yıl (aralığı: 26–90) idi ve %39,5’i kadındı. Tümörlerin çoğunluğu sol kolonda yerleşmişti ( n = 100). Ortalama tümör çapı 4,7 cm idi (aralığı 1,2–13). 124 hastanın klinikopatolojik özellikleri Tablo 1’de verilmiştir .
PD-L1, 15 (%12,1) vakada pozitif ve 109 (%87,9) vakada negatifti [ Şekil 1 a-b]. Vakaların 75’inde (%60,5) PD-L1 bağışıklık hücrelerinde pozitif ve 49’unda (%39,5) negatifti. PD-L2, 52 (%42) vakada pozitif ve 72 (%58) vakada negatifti. PD-L2 boyama yoğunluğu değerlendirildiğinde, 44 hastada hafif boyama, 22 hastada orta düzeyde boyama ve 20 hastada şiddetli boyama vardı [ Şekil 1 c].
Şekil 1:(ac): (a) Medüller özelliklere sahip bir tümörde PD-L1 boyama (×200) (b) Orta derecede farklılaşmış tümörde PD-L1 boyama (×200), (c) Tümör hücrelerinde şiddetli PD-L2 boyama (×200)
Çevre tümör dokusu CD8 ile değerlendirildiğinde 53 (%42,7) olguda lenfosit yüksek düzeyde, 71 (%57,3) olguda lenfosit düşük düzeyde bulundu. TIL’li CD8 12 (%9,7) olguda yüksek düzeyde, 112 (%90,3) olguda ise düşük düzeydeydi.
Tümör hücrelerindeki PD-L1 ekspresyonu kadınlarda ( P < 0,05) ve medüller morfolojiye sahip tümörlerde ( P < 0,05) anlamlı olarak daha yüksekti. T evresi değerlendirildiğinde, PD-L1 ekspresyonu erken evre tümörlerde (T1 ve T2) tümörü çevreleyen bağışıklık hücrelerinde anlamlı olarak daha yüksekti ( P < 0,05) [ Şekil 2 a]. PD-L1 pozitiflik oranı da erken T evre tümörlerindeki tümör hücrelerinde daha yüksekti. Ancak istatistiksel olarak anlamlı bir sonuca ulaşılamadı ( P > 0,05). PD-L1 ekspresyonu işaretli halka hücreli karsinom ve musinöz karsinomda gözlenmedi. Medüller morfolojiye sahip hastalarda, PD-L1 ekspresyonu tümörü çevreleyen bağışıklık hücrelerinde daha yüksek bulundu ancak istatistiksel olarak anlamlı bir sonuca ulaşılamadı. Tümör ve çevresindeki bağışıklık hücrelerinde PD-L1 ekspresyonu ile yaş, tümör yerleşimi, tümör boyutu, histolojik derece, T evresi, N evresi, perinöral invazyon, küçük ve büyük damar invazyonu ve nekroz varlığı arasında istatistiksel olarak anlamlı bir fark bulunmadı ( P > 0,05) [ Tablo 2 ]. CD8 pozitif T hücrelerinin PD-L1 pozitif tümörlerde adaptif bağışıklık direnci ile ilişkili olduğu düşünüldüğünden, çalışmamızda tümör stromasındaki CD8 pozitif lenfosit infiltrasyonu değerlendirildi. PD-L1 pozitifliğinin, CD8 pozitif peritümoral lenfosit ve TIL bulunan vakalarda tümörü çevreleyen bağışıklık hücrelerinde anlamlı derecede daha yüksek olduğu bulundu ( P < 0,05) [ Tablo 3 ]. PD-L1 pozitif tümör hücreleri ile CD8 pozitif peritümoral ve TIL arasında anlamlı bir korelasyon bulunmadı [ Şekil 2 b ].
Şekil 2:(a ve b): (a) PD-L1 pozitif peritümoral ve tümör infiltre eden lenfositlere sahip kolorektal kanser (×200), (b) CD8 pozitif peritümoral ve tümör infiltre eden lenfositlere sahip kolorektal kanser (×200)Tablo 2:Tümör hücrelerinde PD-L1 ekspresyonu ile klinikopatolojik parametreler arasındaki karşılaştırmaTablo 3:Bağışıklık hücresi tarafından PD-L1 ekspresyonunun ve CD8 infiltrasyonu arasındaki ilişki
Medüller morfolojiye sahip tümörlerde PD-L2 ekspresyonu gözlenmedi. İstatistiksel analizde, PD-L2 boyama ile medüller morfoloji arasında ters ve anlamlı bir ilişki vardı ( P < 0,05). PD-L1 pozitif boyama vakalarının üçü PD-L2 ile güçlü bir şekilde boyanmıştı, biri orta derecede, 6’sı zayıf ve 5’i negatifti. 12 vakanın 5’inde CD8 yüksek TIL boyaması PD-L2 pozitifti ve çevreleyen tümör lenfositleri yüksek seviyede olan 53 vakanın 25’inde PD-L2 pozitifti. PD-L2 ekspresyonu ile CD8 boyaması ve diğer klinikopatolojik parametreler arasında istatistiksel olarak anlamlı bir ilişki yoktu [ Tablo 4 ].
Tablo 4:PD-L2 ekspresyonu ile klinikopatolojik parametreler arasındaki ilişki
CD8-yüksek TIL boyanması olan 12 vakanın 10’unda, tümörün etrafında yüksek seviyede lenfositler de vardı. Bu iki grup arasında istatistiksel olarak anlamlı bir ilişki vardı ( P = 0,007). CD8 ile yoğun TIL boyanması, sağ kolon tümörlerinde ve medüller morfolojiye sahip tümörlerde anlamlı derecede daha yüksekti ( P < 0,05).
Hayatta kalma analizi
Takip süresi 56 ± 3,9 ay (aralığı 49-64 ay) olarak belirlendi. Tüm hastaların ortalama nükssüz sağ kalımı 32 ± 3,7 ay (aralığı 24-39 ay) idi. Hasta kohortunun tek değişkenli analizi, PD-L1 pozitif bağışıklık hücrelerini ( P 0,025) nükssüz sağ kalım için önemli bir öngörücü olarak tanımladı [ Tablo 5 ]. Çok değişkenli analizde, PD-L1 pozitif bağışıklık hücrelerinin nükssüz sağ kalımla ilişkili bağımsız faktörler olduğu gösterildi (tehlike oranı 0,56, P 0,03) [ Tablo 6 ]. PD-L1 pozitif ve negatif tümör hücreli olgularda, nükssüz sağ kalım ve takip sürelerinin medyan (ay) değerleri sırasıyla 7 (0-29) ve 30 (5-79), 7 (0-30) ve 13 (1-49) olarak bulundu ( P = 0,471, P = 0,724). PD-L1 pozitif ve negatif immün hücreli olgularda medyan (ay) nükssüz sağ kalım ve takip değerleri sırasıyla 11 (1-26) ve 19 (4-70), 16 (0-42) ve 42 (17-72) olarak bulundu ( P = 0,332, P = 0,165). İstatistiksel değerlendirmede, her ikisinde de anlamlılık bulunmadı. PD-L2 pozitif ve negatif tümör hücreli olgularda nükssüz sağ kalım ve takip sürelerinin medyan (ay) değerleri sırasıyla 7 (0-24) ve 40 (7-74), 6 (0-41) ve 40 (7-84) olarak bulundu. PD-L2 pozitif ve negatif immün hücreli olgularda medyan (ay) nükssüz sağ kalım ve takip değerleri sırasıyla 8 (0-31) ve 31 (8-72), 10 (0-56) ve 21 (4-86) olarak bulundu. Tümör hücrelerinde ( P = 0,543, P = 0,579) ve immün hücrelerde ( P = 0,863, P = 0,961) PD-L2 pozitifliği nükssüz sağ kalım ve takip süreleriyle karşılaştırıldığında anlamlı bir fark bulunmadı. Peritumoral/intratumoral CD8 boyanması ile nükssüz sağkalım ( P = 0.672, P = 0.952) ve takip süreleri ( P = 0.147, P = 0.551) arasında istatistiksel olarak anlamlı bir ilişki saptanmadı. Perinöral invazyon olan olgular ile takip süreleri arasında istatistiksel olarak anlamlı fark bulundu ( P = 0.006). Diğer klinikopatolojik parametrelerle nükssüz sağkalım ve takip süreleri karşılaştırıldığında istatistiksel olarak anlamlı bir fark bulunmadı.
Tablo 5:Klinikopatolojik özellikler ile RFS arasındaki ilişkiTablo 6:Klinikopatolojik özellikler ile RFS arasındaki ilişki
TARTIŞMA
Kolorektal kanser (CRC), dünya çapında yeni teşhis edilen kanser vakalarının yaklaşık %9,7’sini oluşturur. Dünyada erkeklerde ve kadınlarda en sık görülen üçüncü kanser türüdür. [ 1 , 2 ] Kolorektal kanserlerin dağılımı coğrafi bölgelere göre değişir. Kuzeybatı Avrupa, Kuzey Amerika ve Anglosakson bölgelerinde yaygındır ve Afrika, Asya ve Güney Amerika’nın bazı bölgelerinde daha az yaygındır. Her etnik grupta farklılık gösterebilmesine rağmen erkeklerde kadınlardan daha yaygındır. [ 8 , 12 , 13 ] Çalışmamızda vakaların %60,5’i erkek ve %39,5’i kadındı. Kolorektal kanserlerin insidansı yaşla birlikte artar. Ortalama sunum yaşı 62’dir. 40 yaşın altındaki vakalar kalıtsal kolorektal kanser sendromlarından birine sahip olma eğilimindedir. [ 14 ] Çalışmamızda ortalama yaş 64,9’du.
Tüm CRC’lerin yaklaşık %50’si rektum ve sigmoidde yer alır. Ancak sağ kolonun insidansı son on yılda artmıştır. Literatürde, son 30 yılda beyaz erkeklerde sağ kolon tümörlerinin %22’den %38’e çıktığını belirten yayınlar bulunmaktadır.[ 15 ] Çalışmamızda, vakaların %80,6’sı sol kolonda, %19,4’ü ise sağ kolonda yerleşmiştir. Sağ kolon tümörlerinin 13’ü (%54,2) kadın, 11’i (%45,8) erkekti.
Lenfositik infiltrasyonun CRC’lerde prognostik önemi vardır. Çok sayıda çalışma, CD8 pozitif sitotoksik T hücreleri ile infiltre olmuş tümörlerde daha iyi bir prognoz olduğunu bildirmiştir. Bu, T hücresi aracılı anti-tümör bağışıklık tepkisinin prognoz üzerindeki etkisini yansıtır. [ 16 , 17 ] Lenfositik infiltrasyon, mikrosatelit instabilitesi yüksek (MSI-H) tümörlerde önemli ölçüde daha belirgindir. [ 18 ] Tümör hücreleri, T hücresi aracılı bağışıklık tepkisinden kaçmak için bağışıklık sisteminin negatif düzenleyicileri olan bazı molekülleri ifade eder. [ 19 ] PD-1, T hücresi aracılı tepkiyi baskılayan bağışıklık kontrol noktası moleküllerinden biridir. PD-1’in PD-L1 ve PD-L2 olmak üzere iki liganı vardır. PD-1’in PD-L1 ve PD-L2 ile bağlanması, T hücrelerinin aktivasyonunu inhibe ederek bağışıklık sistemini baskılar. PD-L1 ve PD-L2 ifade eden tümörler sitotoksik T hücrelerinden kaçar ve tümörün ilerlemesini engeller.[ 20 ]
PD-1, CD28 ailesinin bir üyesidir ve T hücrelerinde ifade edilir. PD-L’lerin ifadesi değişkendir. PD-L1, PD-L2’den daha yaygın olarak ifade edilir. PD-L1, B hücreleri, dendritik hücreler, makrofajlar, T hücreleri ve endotel hücreleri, pankreas adacık hücreleri, astrositler, keratinositler gibi hematopoietik olmayan hücrelerde ifade edilir. PD-L2, dendritik hücreler, makrofajlar ve mast hücrelerinde ifade edilir.[ 4 , 20 ] Tümör hücreleri hem PD-L1 hem de PD-L2 ifade edebilir.[ 21 ]
Melanom, küçük hücreli olmayan akciğer karsinomu, böbrek hücreli karsinomu, meme kanseri, özofageal kanser, mide kanseri ve lenfomaların PD-L1 ekspresyonu rapor edilmiş ve zayıf klinik seyirle ilişkilendirilmiştir. [ 22 , 23 , 24 , 25 , 26 ] PD-L1 ekspresyonu çok çeşitli katı tümörlerde ve hematolojik malignitelerde zayıf prognozla ilişkilendirilmiştir ve bu da immünomodülatör eksende tedaviyi cazip hale getirir. [ 27 ] Kanser immünoterapisinde, PD-1 ve PD-L1’i spesifik olarak bloke eden antikorlar vardır. Ancak, PD-L2’nin bağışıklık yanıtını düzenlemedeki rolü ve önemi henüz net değildir. Bu nedenle, kanser immünoterapisi için henüz net bir hedef yoktur. [ 28 ]
CRC’de Wang ve diğerleri [ 29 ] tümör hücrelerinde %8 oranında PD-L1 ekspresyonu ve tümöre sızan bağışıklık hücrelerinde %21 oranında PD-L1 ekspresyonu buldular. Vakaların %4’ünde hem tümör hücrelerinde hem de tümöre sızan bağışıklık hücrelerinde PD-L1 ekspresyonu gözlemlediler. Tümöre sızan bağışıklık hücrelerinde PD-L1’in boyanması hastalıksız sağ kalımla ilişkilendirildi. PD-L1’in tümör boyanması ile klinikopatolojik veriler arasında bir korelasyon bulunamadı.
Gasser ve ark . [ 9 ], primer CRC’lerdeki tümör hücrelerinde PD-L1 ve PD-L2 ile membranöz ve sitoplazmik boyama gözlemlediler. Vakaların %42,2’sinde PD-L1 ekspresyonu, %41,4’ünde PD-L2 ekspresyonu ve %24,1’inde hem PD-L1 hem de PD-L2 ekspresyonu buldular. PD-L1 ve PD-L2 ekspresyonu, ileri evre hastalıktaki tümör hücrelerinde önemli ölçüde daha yüksekti. Evreden bağımsız olarak, PD-L1 ve PD-L2 ekspresyonunu eksprese eden tümörlerin sağkalımı önemli ölçüde daha kötüydü. PD-L1 ve PD-L2 ekspresyonunu sağkalımda bağımsız prognostik faktörler olarak buldular. PD-L1 eksprese eden tümörlerin sağkalımı, PD-L2 pozitif tümörlerden daha kötüydü. Ek olarak, hem PD-L1 hem de PD-L2 eksprese eden tümörlerde sağkalım, her iki ligandı da eksprese etmeyenlere göre önemli ölçüde daha kötüydü.
Evre 3 CRC vakalarında, Koganemaru ve diğerleri [ 11 ] tümör hücrelerinin %8,1’inde ve tümörü infiltre eden mononükleer hücrelerin %15,3’ünde PD-L1 ekspresyonu gözlemlediler. Hastalıksız sağ kalımın tümör hücrelerinde PD-L1 ekspresyonunda önemli ölçüde daha kısa olduğunu buldular. Buna karşılık, hastalıksız sağ kalım, tümörü infiltre eden mononükleer hücrelerde PD-L1 ekspresyonu olan vakalarda önemli ölçüde daha uzundu.
Lee ve arkadaşları [ 30 ], PD-1 ve PD-L1 ekspresyonunun MSI-H’li hastalarda anlamlı derecede daha yüksek olduğunu buldular. Buna uygun olarak, bu çalışmada, medullar özellikler, yüksek TIL ve yüksek tümör çevresi lenfositik agregasyonunun da PD-L1 ekspresyonuyla anlamlı derecede ilişkili olduğunu buldular. Rosenbaum ve arkadaşları [ 12 ], PD-L1 ekspresyonunun kadın, medullar fenotip, yoğun TIL ve MSI-H vakalarında anlamlı derecede daha yüksek olduğunu buldular. Diğer tümör tiplerinden farklı olarak, CRC PD-1 ve PD-L1 blokajına düşük oranda yanıt verir. [ 6 ] Le ve arkadaşları [ 31 ], metastatik CRC’li hastalarda pembrolizumabın klinik etkilerini araştıran çalışmalarında, MSI-H genotipli hastaların bu tedaviye anlamlı şekilde yanıt verdiğini bildirdiler. Bu çalışmanın sonucunda pembrolizumab tedavisi uygulanacak hastaların seçiminde MMR durumunun belirlenmesinin önemli olduğu vurgulanmıştır.
Masugi ve diğerleri [ 32 ], PD-L2 boyama ile lenfositik reaksiyon ve CRC’lerdeki klinikopatolojik parametreler arasındaki ilişkiyi araştıran çalışmalarında, PD-L2 ekspresyonunun kadınlarda, iyi ve orta derecede farklılaşmış tümörlerde ve yüksek PD-L1 ekspresyonu gösteren tümörlerde önemli ölçüde daha yüksek olduğunu buldular. PD-L2 ekspresyonunun Crohn benzeri lenfoid reaksiyonlu hastalarda önemli ölçüde daha düşük olduğunu buldular. Tümör hücrelerindeki PD-L2 ekspresyonunun lenfoid yanıtın gelişimini engellediğini öne sürdüler.
Çalışmamızda tümör hücrelerinin %12,1’inde PD-L1 ekspresyonu, tümörü çevreleyen immün hücrelerin ise %60,5’inde PD-L2 ekspresyonu gözlendi. Tümör hücrelerinde PD-L2 ekspresyonu olguların %42’sinde saptandı. Literatürle benzer şekilde tümör hücrelerinde PD-L1 ekspresyonu kadınlarda ve medüller morfolojiye sahip hastalarda anlamlı olarak yüksekti ( P < 0,05). Medüller morfolojiye sahip tümörlerde PD-L2 ekspresyonu gözlenmezken, PD-L2 ile medüller morfoloji arasında istatistiksel olarak ters ilişki bulundu ( P < 0,05). PD-L1 pozitif boyanma olgularının 3’ü PD-L2 ile güçlü boyanmıştı. CD8-yüksek TIL boyanması olan 12 olgunun 5’inde PD-L2 pozitif, çevresindeki tümör lenfositleri yüksek olan 53 olgunun 25’inde PD-L2 pozitifti. Bu çalışmada tümör çevresindeki immün hücrelerde ve erken T evresi (T1 ve T2) vakalarda PD-L1 ekspresyonu arasında istatistiksel olarak anlamlı ilişki bulunmuştur ( P < 0,05). Ayrıca tümör çevresinde yoğun CD8 ve TIL ve lenfosit bulunan vakalarda tümör çevresindeki immün hücrelerde PD-L1 ekspresyonu anlamlı olarak daha yüksek bulunmuştur ( P < 0,05). Literatürden farklı olarak tümör hücrelerinde PD-L1 ekspresyonu erken T evresi tümörlerde daha fazla gözlenmiş ancak istatistiksel olarak anlamlı bir fark bulunamamıştır. Literatürle benzer şekilde, PD-L1 pozitif tümör infiltre eden lenfositlerin aslında nüks içermeyen sağkalımı etkileyebileceğini gösterdik.
Bu çalışmada, PD-L1, CRC’de hem tümör hem de bağışıklık hücrelerinde ifade edildi. Yüksek CD8+ T hücreli hastalarda yüksek PD-L1 ifadesi, adaptif bağışıklık direnci mekanizmasıyla ilişkiyi destekler. Kadın cinsiyet, medüller morfoloji, tümörün içinde ve çevresinde yoğun lenfositik infiltrasyon, MSI-H tümörlerinde ortak özelliklerdir. [ 21 ] Çalışmamızda, PD-L1 ifadesinin kadınlarda, medüller morfolojide ve TIL ve tümör açısından zengin lenfositik infiltrasyonda önemli ölçüde daha yüksek olduğu bulundu. Bu bulgular, literatürde bildirilen CRC’lerdeki PD-L1 ekspresyonu ile mikrosatelit instabilitesi arasındaki ilişkiyi desteklemektedir. Medüller morfolojili tümörlerde ve kadın cinsiyette PD-L1’in önemli ekspresyonu, hasta seçiminde yararlı olabileceğini düşündürmektedir.
Finansal destek ve sponsorluk
Çalışmamız Eskişehir Osmangazi Üniversitesi Bilimsel Araştırma Proje Birimi tarafından desteklenmiştir.
Çıkar çatışmaları
Herhangi bir çıkar çatışması bulunmamaktadır.
KAYNAKLAR
1. Siegel RL, Miller KD, Fuchs HE, Jemal A. Kanser istatistikleri, 2021 CA Cancer J Clin. 2021;71:7–33
2. Siegel RL, Miller KD, Jemal A. Kanser istatistikleri, 2018 CA Cancer J Clin. 2018;68:7–30
3. Pagès F, Berger A, Camus M, Sanchez-Cabo F, Costes A, Molidor R, ve diğerleri Etkili hafıza T hücreleri, erken metastaz ve kolorektal kanserlerde sağ kalım N Engl J Med. 2005;353:2654–66
4. Keir ME, Francisco LM, Sharpe AH. PD-1 ve T-hücre bağışıklığındaki ligandları Curr Opin Immunol. 2007;19:309–14
5. Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ, Topalian SL, Hwu P, ve diğerleri İleri kanserli hastalarda anti-PD-L1 antikorunun güvenliği ve aktivitesi New Engl J Med. 2012;366:2455–65
6. Yoneda K, Imanishi N, Ichiki Y, Tanaka F. Küçük hücreli olmayan akciğer kanserinde (KHDAK) immün kontrol noktası inhibitörleri (ICI’ler) J UOEH. 2018;40:173–89
7. Wang X, Teng F, Kong L, Yu J. İnsan kanserlerinde PD-L1 ekspresyonu ve klinik sonuçlarla ilişkisi Onco Targets Ther. 2016;9:5023–39
8. Murphy G, Devesa SS, Cross AJ, Inskip PD, McGlynn KA, Cook MB. Anatomik alt bölgeye, ırka ve yaşa göre kolorektal kanser insidansındaki cinsiyet farklılıkları Int J Cancer. 2011;128:1668–75
9. Grimm M, Gasser M, Koenigshausen M, Stein C, Lutz J, Krol S, ve diğerleri İnsan kolorektal kanserinde programlanmış ölüm-1 ligand-1 ve programlanmış ölüm-1 ligand-2 ekspresyonunun klinik önemi ve terapötik potansiyeli J Clin Oncol. 2008;26:15005
10. Sridharan V, Gjini E, Liao X, Chau NG, Haddad RI, Severgnini M, ve diğerleri Adenoid kistik karsinomun bağışıklık profili: PD-L2 ekspresyonu ve tümör infiltre eden lenfositlerle ilişkileri Cancer Immunol Res. 2016;4:679–87
11. Koganemaru S, Inoshita N, Miura Y, Miyama Y, Fukui Y, Ozaki Y, ve diğerleri Evre III kolorektal kanserli hastalarda programlanmış ölüm ligandı 1 ekspresyonunun prognostik değeri Cancer Sci. 2017;108:853–8
12. Rosenbaum MW, Bledsoe JR, Morales-Oyarvide V, Huynh TG, Mino-Kenudson M. Kolorektal kanserdeki PD-L1 ekspresyonu mikrosatelit instabilitesi, BRAF mutasyonu, medüller morfoloji ve sitotoksik tümör infiltre eden lenfositlerle ilişkilidir Mod Pathol. 2016;29:1104–12
14. Yantiss RK, Goodarzi M, Zhou XK, Rennert H, Pirog EC, Banner BF, ve diğerleri Erken başlangıçlı kolorektal karsinomun klinik, patolojik ve moleküler özellikleri Am J Surg Pathol. 2009;33:572–82
15. Cucino C, Buchner AM, Sonnenberg A. Kolon Rektum Kolorektal kanserinin sağa doğru kaymasının devam etmesi. 2002;45:1035–40
16. Naito Y, Saito K, Shiiba K, Ohuchi A, Saigenji K, Nagura H, ve diğerleri İnsan kolorektal kanserinde prognostik bir faktör olarak kanser hücre yuvalarına sızan CD8+T hücreleri Cancer Res. 1998;58:3491–4
17. Menon AG, Janssen-van Rhijn CM, Morreau H, Putter H, Tollenaar RA, van de Velde CJ, ve diğerleri Bağışıklık sistemi ve kolorektal kanserde prognoz: Ayrıntılı bir immünohistokimyasal analiz Lab Invest. 2004;84:493–501
22. Thompson RH, Dong H, Lohse CM, Leibovich BC, Blute ML, Cheville JC ve diğerleri PD-1 tümör infiltre eden bağışıklık hücreleri tarafından ifade edilir ve böbrek hücreli karsinomlu hastalarda kötü sonuçlarla ilişkilidir Clin Cancer Res. 2007;13:1757–6
23. Konishi J, Yamazaki K, Azuma M, Kinoshita I, Dosaka-Akita H, Nishimura M. Küçük hücreli olmayan akciğer kanseri hücrelerinde B7-H1 ekspresyonu ve tümör infiltre eden lenfositler ve PD-1 ekspresyonu ile ilişkisi Clin Cancer Res. 2004;10:5094–100
24. Muenst S, Schaerli A, Gao F, Däster S, Trella E, Droeser R, ve diğerleri Programlanmış ölüm ligandı 1’in (PD-L1) ekspresyonu insan meme kanserinde kötü prognozla ilişkilidir. Breast Cancer Res Treat. 2014;146:15–24
25. Chen L, Deng H, Lu M, Xu B, Wang Q, Jiang J, ve diğerleri B7-H1 ekspresyonu tümör invazyonu ile ilişkilidir ve insan özofagus kanseri hastalarının sağ kalımını tahmin eder Int J Clin Exp Pathol. 2014;7:6015–
26. Hou J, Yu Z, Xiang R, Li C, Wang L, Chen S, ve diğerleri Mide kanseri tümör dokularında FOXP3+ düzenleyici T hücrelerinin infiltrasyonu ile B7-H1 ekspresyonu arasındaki korelasyon Exp Mol Pathol. 2014;96:284–91
27. Chen DS, Irving BA, Hodi FS. Moleküler yollar: Yeni nesil immünoterapi: Programlanmış ölüm-ligand 1 ve programlanmış ölüm-1’i inhibe etme Clin Cancer Res. 2012;18:6580–7
28. Rozali EN, Hato SV, Robinson BW, Lake RA, Lesterhuis WJ. Kanser kaynaklı bağışıklık baskılanmasında programlanmış ölüm ligandı 2 Clin Dev Immunol. 2012:2012
29. Wang L, Ren F, Wang Q, Baldridge LA, Monn MF, Fisher KW, ve diğerleri Kolorektal kanserlerde programlanmış ölüm ligandı 1 (PD-L1) immünohistokimyasal ekspresyonunun önemi Mol Diagn Ther. 2016;20:175–81
30. Lee LH, Cavalcanti MS, Segal NH, Hechtman JF, Weiser MR, Smith JJ, ve diğerleri Kolorektal karsinomda PD-1 ve PD-L1 ekspresyonunun kalıpları ve prognozla ilişkisi Mod Pathol. 2016;29:1433–42
31. Le DT, Uram JN, Wang H, Bartlett BR, Kemberling H, Eyring AD, ve diğerleri Uyumsuzluk onarım eksikliği olan tümörlerde PD-1 blokajı N Engl J Med. 2015;372:2509–20
32. Masugi Y, Nishihara R, Hamada T, Song M, Da Silva A, Kosumi K, ve diğerleri Tümör PDCD1LG2 (PD-L2) ekspresyonu ve kolorektal kansere karşı lenfositik reaksiyon Cancer Immunol Res. 2017;5:1046–55
Magnapinna Mürekkep Balığını (bigfin kalamarı) Gözlemlemek için yorumlar kapalı
Bigfin kalamarları, kendilerine özgü bir morfolojiye sahip olan, nadiren görülen bir kafadanbacaklılar grubudur. Magnapinnidae familyasına yerleştirilirler.
Ailenin yalnızca larva, paralarval ve genç örnekleri görülmüştür. Zamanla, Magnapinnalara benzer bir morfolojiye sahip olduğu görülen ve “uzun kollu kalamar ” lakaplı olan bazı hayvanların birkaç videosu çekilmiştir.
Ancak bu canlıların yetişkin örneklerinden hiçbiri yakalanmadığı veya örneklenmediği için, bunların aynı cinsten mi yoksa sadece uzak akraba mı olduğu bilinmemektedir.
Bigfin kalamarının kolları ve dokunaçları son derece uzundur. Bu kolların 4 ila 8 metre uzunluğunda olduğu düşünülmektedir. Bu uzantılar vücuda dikey durarak canlının dirseklerini oluşturmaktadırlar.
Bilim insanları, bigfin kalamarının kollarını ve dokunaçlarını deniz tabanı boyunca sürükleyerek yerden yenilebilir organizmaları kaptığını ve beslendiğini düşünmektedirler.



_(14802506383).jpg)



/storage.evrimagaci.org%2Fcontent%2F0100116a-b991-4027-b603-e6d3fc5db54e.jpeg)
/storage.evrimagaci.org%2Fold%2Fcontent_media%2Fd1a8032e99451f2f261e178218df8a3d.jpg)
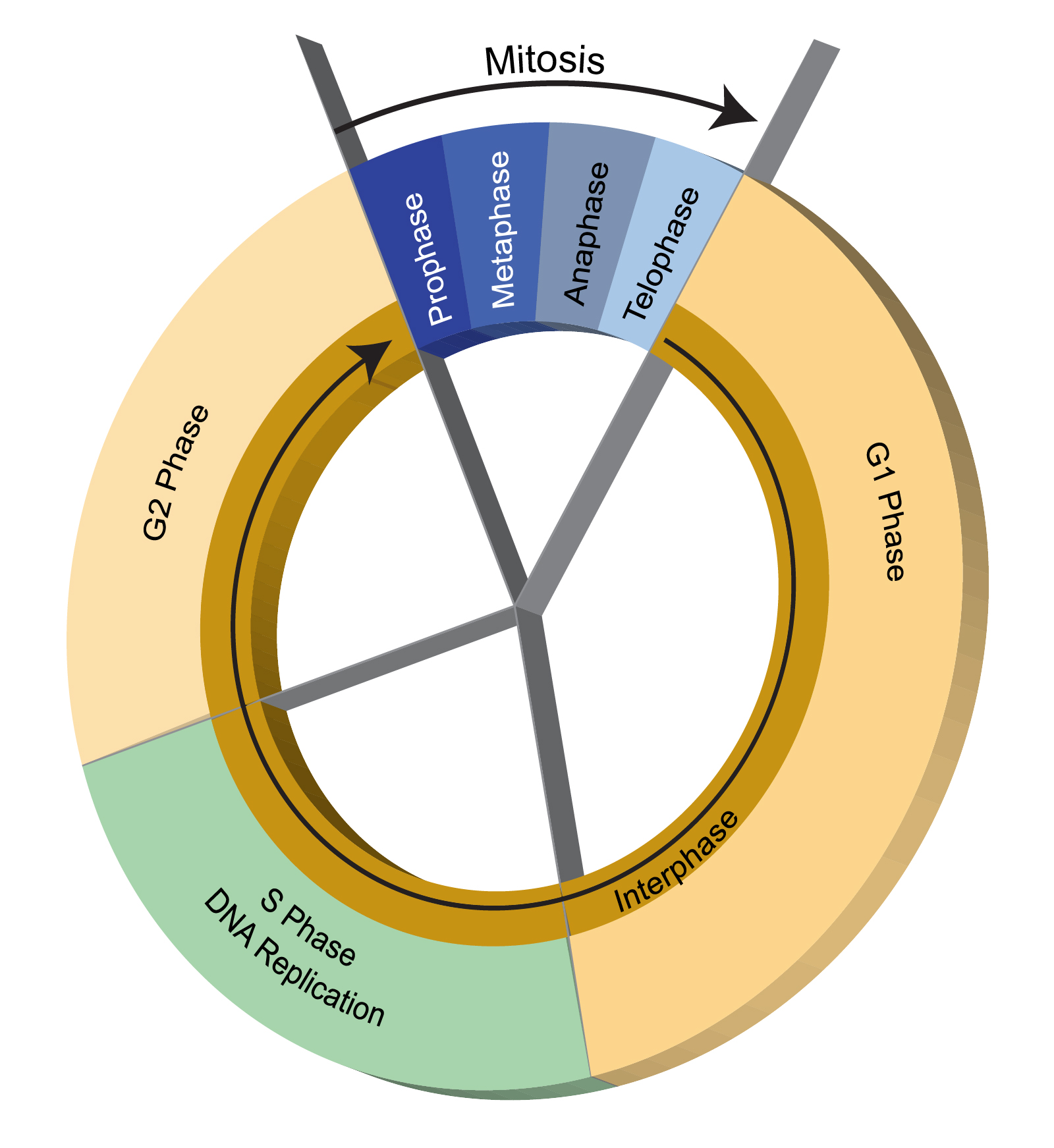
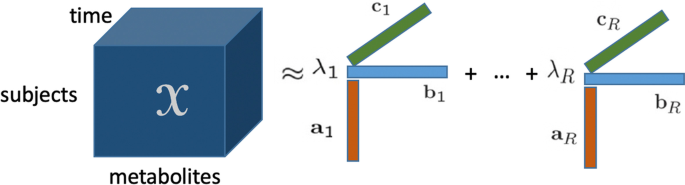
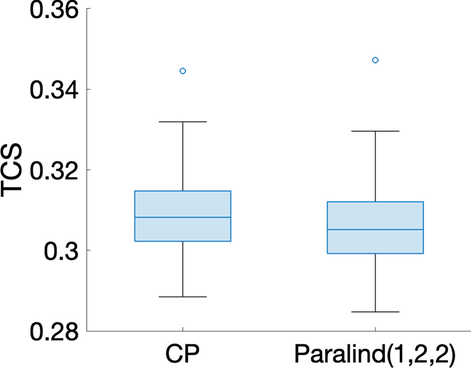
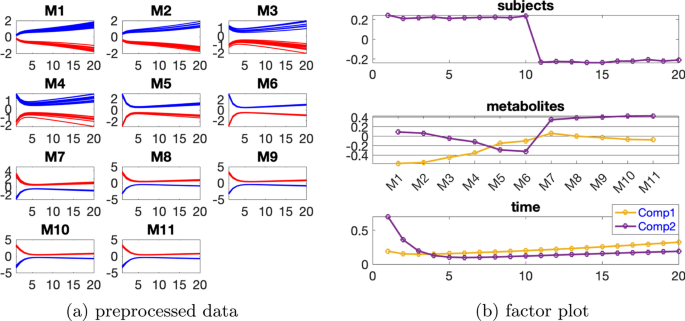
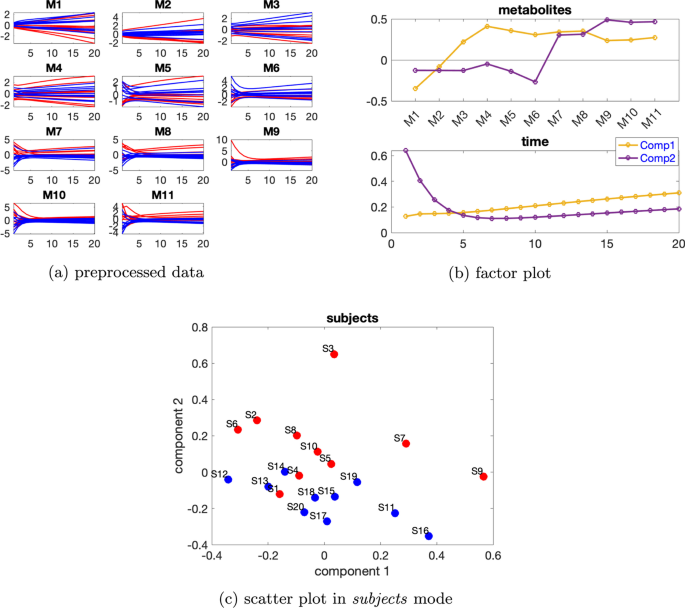
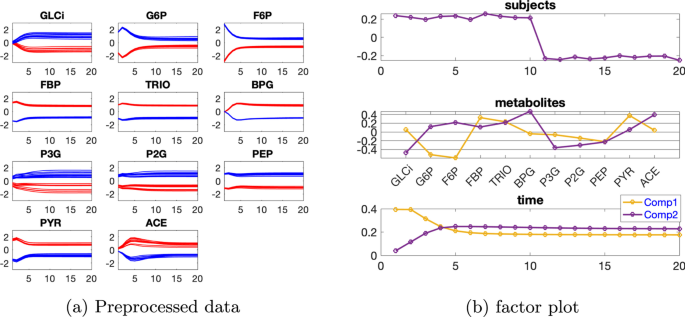
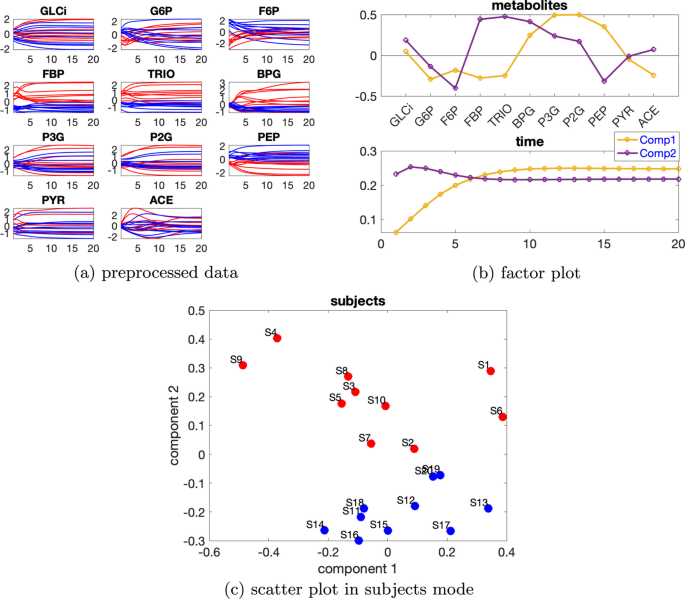
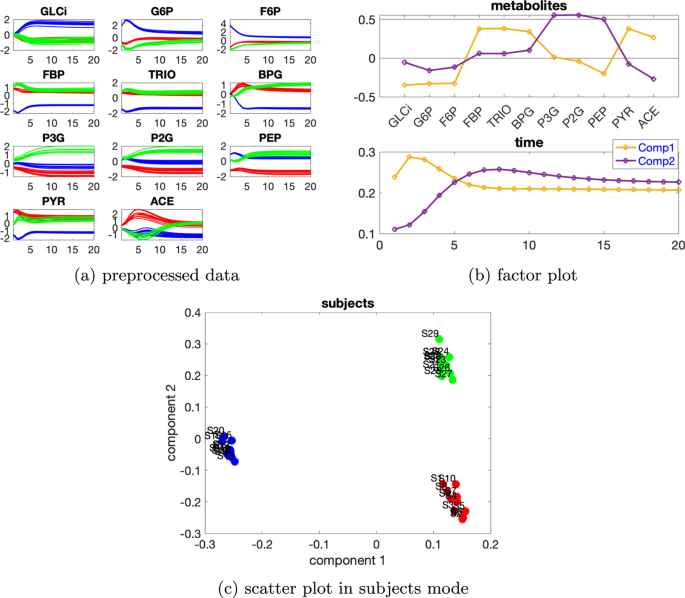
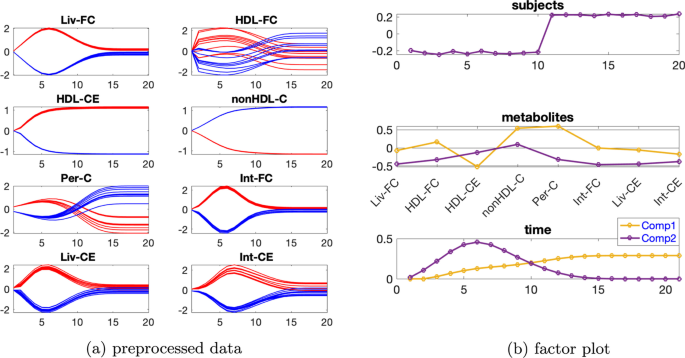
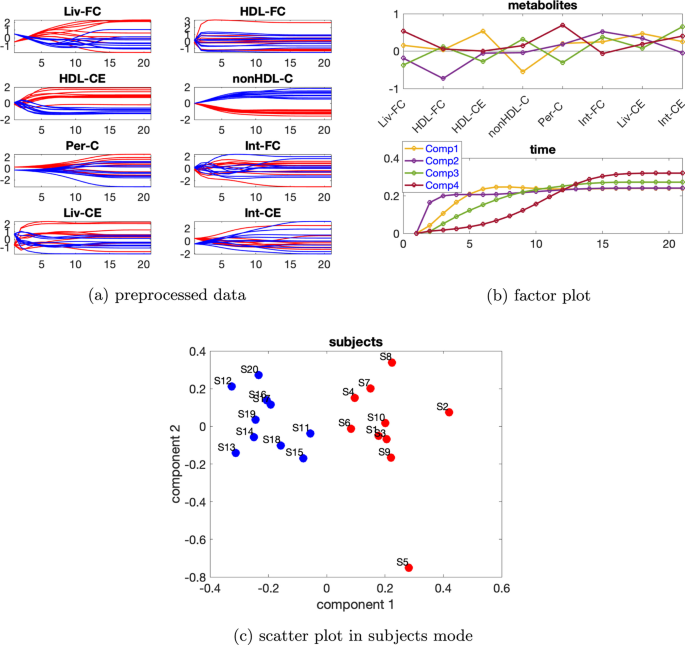
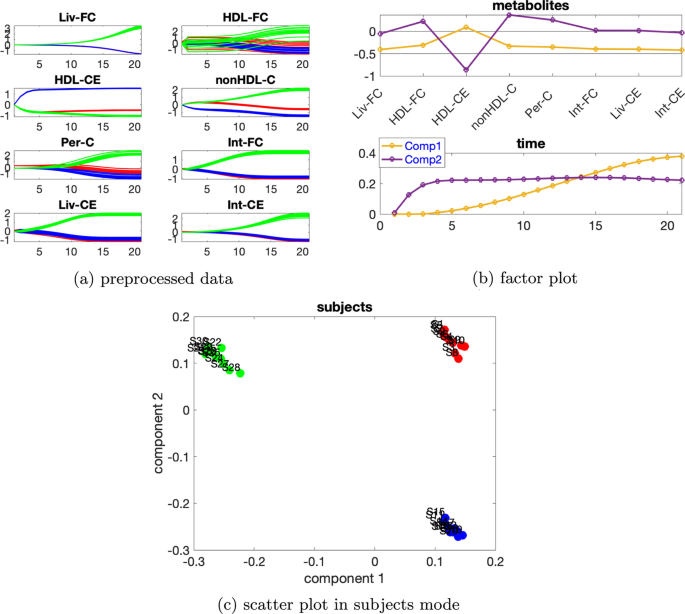


/storage.evrimagaci.org%2Fold%2Fcontent_media%2Fd4f1c3600ff320769ec0eb82078ce989.png)








