Rawsamble: Hash tabanlı bir tohumlama mekanizması kullanarak ham nanopore sinyallerini üst üste bindirme ve birleştirme için yorumlar kapalı
Ham nanopore sinyal analizi, sinyalleri bazlara çevirmeden (yani baz çağrısı yapmadan) hızlı ve kaynak açısından verimli analiz sağlamak için genomikte yaygın bir yaklaşımdır. Ancak, çiftler halinde ham sinyal karşılaştırmasında artan gürültüyü ele almak için doğru mekanizmaların eksikliği nedeniyle referans genom bilinmiyorsa mevcut çözümler ham sinyalleri doğrudan yorumlayamaz. Amacımız, referans genom olmadan ham sinyallerin doğrudan analizini sağlamaktır. Bu amaçla, 1) karma tabanlı bir arama mekanizması kullanarak tüm ham sinyal çiftleri arasındaki benzerlik bölgelerini belirleyebilen, tümüne karşı tümü örtüşen olarak bilinen ve 2) bunları sıfırdan genomlar oluşturmak için kullanabilen ilk mekanizma olan Rawsamble’ı öneriyoruz, buna de novo birleştirme denir. Çeşitli boyutlardaki birden fazla genomda yaptığımız kapsamlı değerlendirmeler, Rawsamble’ın temel çağrı (Dorado’nun en hızlı modu) ve bir CPU’da örtüşme (minimap2) için en son teknoloji araçlarını kullanan geleneksel bir genom birleştirme hattına kıyasla önemli bir hızlanma (ortalama 16,36 kat ve 41,59 kata kadar) sağladığını ve en yüksek bellek kullanımını (ortalama 11,73 kat ve 41,99 kata kadar) azalttığını göstermektedir. Rawsamble tarafından üretilen örtüşen çiftlerin %36,57’sinin minimap2 tarafından üretilenlerle aynı olduğunu buluyoruz. Rawsamble’dan gelen örtüşmeleri kullanarak, temel çağrı yapmadan doğrudan ham sinyallerden ilk de novo birleştirmeleri oluşturuyoruz. 2,7 milyon baz uzunluğa kadar (E. coli’nin genom uzunluğunun yarısı) bitişik birleştirme segmentleri (unitig’ler) oluşturabileceğimizi gösteriyoruz. Örtüşmeleri bularak ve de novo birleştirmeleri oluşturarak etkinleştirilebilecek daha önce keşfedilmemiş yönleri belirliyoruz . Ayrıca sonuçlarımızı GitHub sayfamızda tam olarak yeniden üretmek için betikler de sağlıyoruz.
MAKİNA ÖĞRENME YÖNTEMLERİYLE GENOMDİZİLİM VERİLERİNİN ANALİZİ için yorumlar kapalı
Biyoinformatik, biyoloji çalışmalarından elde edilen ham verinin bilgisayarlarla işlenebilir formata getirilmesi, analizi ve uygun formatta saklanması için gerekli tüm yöntemleri bünyesinde toplayan disiplinler arası bir çalışma alanıdır. Bu alanın günümüzdeki popüler çalışma konuları: genom dizileme, genom kapsamında ilişki çıkarımı, dizilim verilerinin analizi, protein sınıflandırma, tür içi ve türler arası çeşitliliklerin tespiti vb. şeklinde özetlenebilir. Bu çalışma konularına bağlı olarak hesaplamalı bilim disiplinlerinden makine öğrenmesi başta olmak üzere sinyal ve görüntü işleme, istatistik ve algoritma tasarımı alanlarındaki yöntemlere sıklıkla başvurulmaktadır.
Genomik Gardiyan (Koruyucu Melek): p53 Geni ve Proteini Nedir, Genlerimizi Nasıl Korur? için yorumlar kapalı
Genomun gardiyanı olarak da tanımlanan p53 proteini, ilk kez 1979 yılında hayvan deneylerinde incelenen tümörlerde belirlenmiş ve ilerleyen 20 yıl boyunca görevi tam olarak anlaşılamamıştır. Kendisini “gen” olarak tabir etsek de aslında TP53 geni tarafından sentezlenen kısa ömürlü bir proteindir. İlk belirlendiğinde (1979) ağırlığı 53 kDa (kilodalton) olarak bulunduğundan “p53” olarak adlandırılmıştır.
p53, temel görevi hücre bölünmesini düzenlemek olan bir transkripsiyon faktördür. Hücre büyümesinin durdurulması, programlanmış hücre ölümü, hücre farklılaşması ve DNA tamir mekanizmasının başlatılması gibi işlevleri ve birçok organizmada kanserin baskılanmasında rolü olan bir proteindir.
“Genomik gardiyan” olarak anılma sebebi, mutant hücre çoğalmasına karşı genomun korunmasında hücrede oluşmuş bir DNA hasarı tamir olana kadar mitoz bölünmeyi durdurmasıdır.
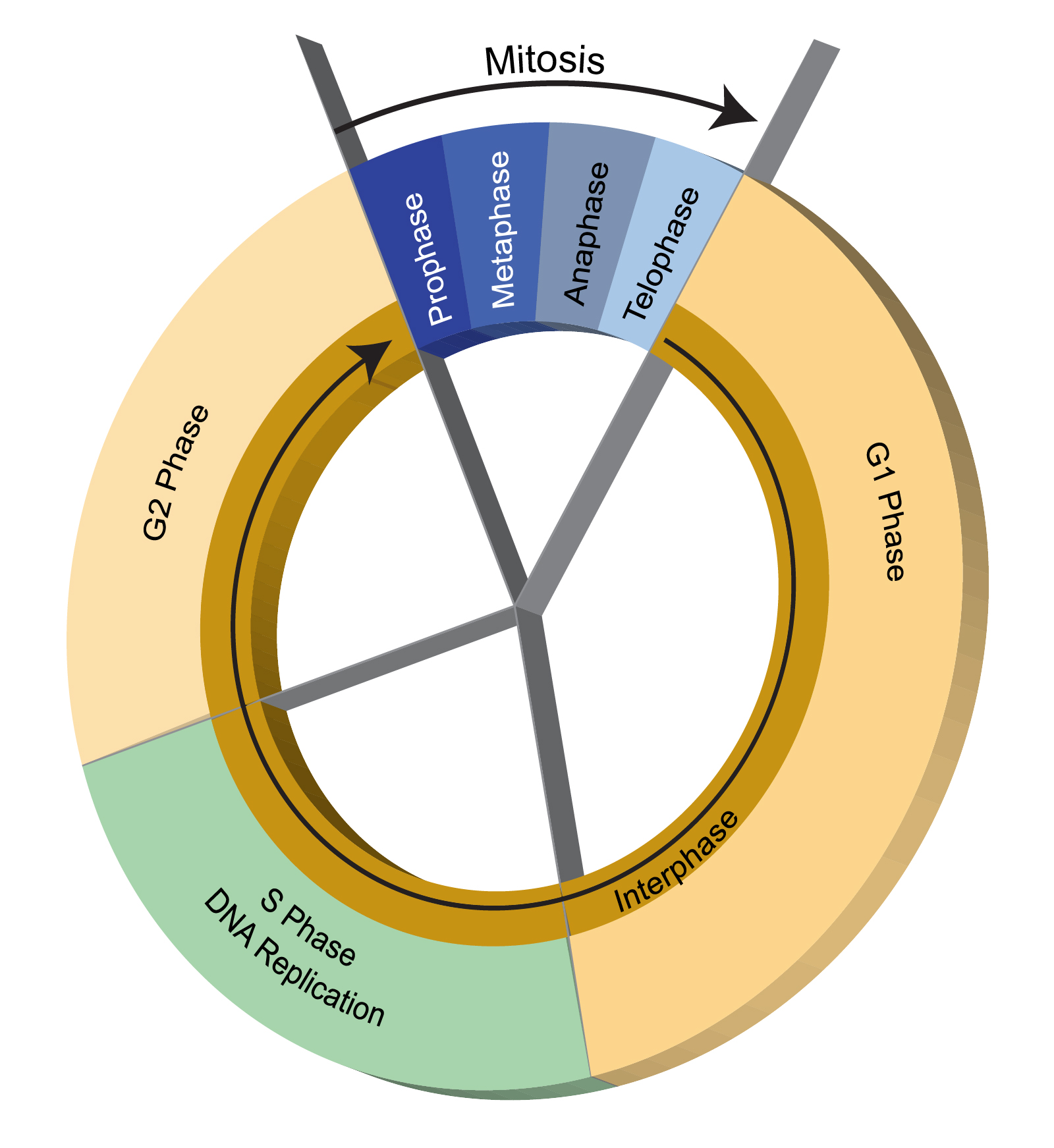
Genotoksik Stres Yokluğunda Hücre Döngüsü
Sürekli bölünen hücrelerde mitozdan sonra döngü (siklus), G1-S-G2 ve tekrardan mitoz şeklinde tekrarlanarak devam eder. Bu süreçte hücre uyarımı ve büyüme meydana gelmekte veya bölünme sinyali gelmediği sürece istirahat fazı olan G0 evresinde durmaktadır.
G1 fazında hücreler kendi çevrelerini kontrol eder sinyalleri alır ve büyümeyi indükler. Bu fazda DNA sentezi (replikasyon) için hazırlık yapılır, RNA ve protein sentezi yapılır. S fazında, DNA sentezlendikten sonra G2 fazında hücre büyümeye devam ederken yine RNA ve protein sentezi gerçekleşir ve hücre mitoza hazırlanır.
Hücre Döngüsü
Hücre siklusunda bir faz tamamlanmadan sonraki faza geçilirse genetik materyal tam ve doğru kopyalanmadığı için hücrede hasar meydana gelebilir. Hücre siklusunda G1-S ve G2-M geçişinde, ayrıca mitosun safhalarından metafazdan anafaza geçişte kontrol noktaları vardır. Bu kontrol noktalarında hücrenin siklusu devam edip etmeyeceği belirlenir.
Genotoksik Stres Olursa Ne Olur?
Radyasyon veya toksinle karşılaşan hücrelerde -genotoksik stres varlığında- DNA’da meydana gelen hasara göre kontrol noktalarında bir sonraki faza geçiş engellenir. Örneğin, DNA sentezindeki bozulmayı algılayıp p53’ü uyaran protein kinaz ailesinden (PI-3) ATM iyonize radyasyon varlığında, ATR ise UV ışın varlığında p53 genini fosforiller. Tümör baskılayıcı p53 geni işlevini kaybederse hücre büyümesinin kontrolü ortadan kalkar ve DNA tamiri olmadan hücre siklusu kontrolsüz devam eder. Oysa normal hücrelerde DNA hasarı olduğunda, p53 genomik kararlılığı sağlar ve hücre siklusunu G1’de inhibe ederek hücreye tamir için zaman kazandırır. Hasar tamir edilemiyorsa hücre planlı olarak intihar eder (hücrenin planlı olarak kendini öldürmesine “apoptoz” denir.).
p53 geninin birçok kanser türünde mutasyona uğramış olduğu ve normalde hücrede oldukça az miktarda bulunmasına rağmen miktarını arttırdığı bilinmektedir. Kanserin basit bir açıklaması olarak kullandığımız “kontrolsüz hücre çoğalması” tanımının altında yatan sebeptir. Hücre çoğalması p53 geni mutasyonu sonucu kontrol edilemez hale gelir. Dokunun asıl hücrelerine benzeyen yada benzemeyen bir çok hücre oluşur. Çoğalan hücre, köken aldığı dokuya ne kadar benziyorsa kanser o kadar “iyi huylu”dur ve buna “iyi diferansiye” deriz.
Kanser teşhisinde p53 miktarı ve yapısını inceleyip kanserin türü, işleyişi, agresifliği hakkında fikir sahibi olmamız mümkündür. Bu diagnostik avantajı haricinde genomik gardiyanımızın ve onu etkileyen moleküllerin günümüzde yoğun olarak inceleniyor olma sebebi, p53’ün bilim insanlarına kanser tedavilerine “Acaba kanserli dokuda sağlıklı p53 geni aktifleştirilerek hücre çoğalması yeniden kontrol altına alınabilir mi?” sorusu ile gelen alternatif bir bakış açısı sunması olmuştur.
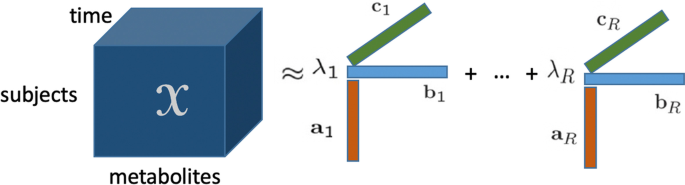
Dinamik metabolomik verilerin analizi, metabolizmadaki altta yatan mekanizmaları anlamamızı geliştirme vaadinde bulunur. Örneğin, bir hastalığın başlangıcından dolayı metabolizmadaki değişiklikleri tespit edebilir. Dinamik veya zamanla çözülmüş metabolomik veriler, bir özne moduna, bir metabolit moduna ve bir zaman moduna göre düzenlenmiş girişlerle üç yönlü bir dizi olarak düzenlenebilir. Bu tür zamanla gelişen çok yönlü veri kümeleri giderek daha fazla toplanırken, bu tür verilerden altta yatan mekanizmaları ve dinamiklerini ortaya çıkarmak zorlu olmaya devam etmektedir. Bu tür veriler için, karmaşıklıklardan biri, çeşitli varyasyon kaynaklarının bir araya gelmesinin varlığıdır: indüklenen varyasyon (deneysel koşullar veya doğuştan gelen hatalar nedeniyle), bireysel varyasyon ve ölçüm hatası. Çok yönlü veri analizi (tensör faktörizasyonları olarak da bilinir), çok yönlü verilerdeki altta yatan kalıpları bulmak için veri madenciliğinde başarıyla kullanılmıştır. Çok yönlü veri analizi yöntemlerinin dinamik metabolomik verilerdeki altta yatan mekanizmaları ortaya çıkarma açısından performansını keşfetmek için, bilinen temel gerçekliğe sahip simüle edilmiş veriler incelenebilir.
Sonuçlar
Artan karmaşıklığa sahip farklı dinamik modellerden, yani basit bir doğrusal sistem, bir maya glikoliz modeli ve bir insan kolesterol modelinden kaynaklanan simüle edilmiş verilere odaklanıyoruz. İndüklenen varyasyonun yanı sıra bireysel varyasyona sahip veriler üretiyoruz. Bu tür dinamik metabolomik verileri analiz etmede çok yönlü veri analizinin avantajlarını ve sınırlamalarını ve farklı varyasyon kaynaklarını çözme kapasitelerini göstermek için sistematik deneyler gerçekleştirilir. Çok yönlü veri analizi yöntemlerinin temel gerçeği bilmenin kolaylaştırdığı yeteneğini anlamak istediğimiz için simülasyonları kullanmayı tercih ediyoruz.
Çözüm
Sayısal deneylerimiz, incelenen dinamik metabolik modellerin giderek karmaşıklaşmasına rağmen, tensör faktörizasyon yöntemleri CANDECOMP/PARAFAC(CP) ve Doğrusal Bağımlılıklara Sahip Paralel Profiller (Paralind)’in varyasyon kaynaklarını çözebileceğini ve böylece altta yatan mekanizmaları ve bunların dinamiklerini ortaya çıkarabileceğini göstermektedir.
Arka plan
Nükleer Manyetik Rezonans (NMR) Spektroskopisi ve Kütle Spektrometrisi (MS) ile gaz kromatografisi (GC) veya sıvı kromatografisi (LC) gibi gelişmiş analitik ölçüm tekniklerinin kullanılabilirliğiyle, biyolojik sistemlerden dinamik veya zamana bağlı (veya uzunlamasına) metabolomik verileri toplamak giderek daha popüler hale geliyor. Bu, bu tür verilerin altta yatan biyolojik süreçleri ve mekanizmaları ortaya çıkarabilme vaadini taşıması nedeniyle daha da popüler hale geliyor. Örnekler, bireylerin sağlık durumlarını araştırmak için meydan okuma testlerinin kullanıldığı metabolizma ve sağlık alanından [ 1 ]; belirli gıda bileşiklerinin metabolik kaderinin incelendiği gıda biliminden [ 2 ]; hastalıkların biyobelirteçlerinin ve hastalık durumlarına erken geçişlerin yakalandığı hastalıkların çalışmasından [ 3 ] vb. verilebilir.
Bahsedilen dinamik metabolomik çalışmalarının temel özellikleri, sınırlı sayıda denekten ölçümlerin alındığı sınırlı sayıda zaman noktası ve farklı varyasyon kaynaklarının üst üste gelmesidir. Farklı varyasyon kaynakları açısından, öncelikle, farklı tedavilerden kaynaklanabilen, örneğin [4]’te ele alınan Qingkailing enjeksiyon grubu veya bir enzimin normalden çok daha düşük aktiviteye sahip olduğu bir hastalıktan kaynaklanabilen, örneğin [ 5 ]’te açıklanan insan mutantları, indüklenmiş varyasyon vardır. İkinci olarak, genellikle oldukça büyük olan bireysel (biyolojik olarak da adlandırılır) varyasyon vardır [ 6 ]. Son olarak, cihaza bağlı olan ve önemli olabilen (kaçınılmaz) ölçüm hatası (teknik hata olarak da adlandırılır) vardır [ 7 ]. Tüm bunlar, bu tür dinamik metabolomik verilerin analizini zorlaştırır.
Bu zorluklar göz önüne alındığında, boyut azaltma yöntemleri, gürültü azaltma (örneğin, ölçüm hatasıyla başa çıkma) ve varyasyonun temel temel kaynaklarını yakalama (dinamik metabolomik verileri analiz etmek için farklı yöntemler hakkında bir inceleme için Smilde ve ark. [ 8 ]’e bakın) için ideal olduklarından umut verici yaklaşımlardır. Boyut azaltma teknikleri, verilerde temelde düşük bir boyutluluk olduğu gerçeğini kullanır ve sözde iki yönlü veriler için bu tür yöntemlerin prototipik örnekleri, örneğin Ana Bileşen Analizi (PCA) ve Ortogonal Kısmi En Küçük Kareler (OPLS), gücünü göstermiştir [ 9 ] ve uzunlamasına metabolomik veri analizi için dinamik olasılıklı PCA’ya uzantılar yapılmıştır [ 10 ]. Veriler özneler , metabolitler ve zaman gibi iki moddan fazla moda sahip olduğunda , verileri iki yönlü bir dizi olarak ele almak yerine çok yönlü bir dizi (daha yüksek dereceli tensör olarak da adlandırılır) oluşturulabilir ve çok yönlü diziler için tensör faktörizasyonları [ 11 , 12 , 13 , 14 ] olarak bilinen boyut azaltma yöntemleri bu tür zamansal verileri analiz etmek için kullanılabilir. Daha önce zamanla değişen metabolomik verileri analiz etmek için kullanılan iki yönlü PCA tabanlı yöntemlerle karşılaştırıldığında, tensör faktörizasyonları tüm modlardaki temel desenleri aynı anda sağlama vaadinde bulunur, örneğin özneler , metabolitler ve zaman modlarındaki desenler. Tensör faktörizasyonları, tartışma izleme [ 15 ], zamansal bağlantı tahmini [ 16 ], veri akışlarının analizi [ 17 ], nörogörüntüleme veri analizi [ 18 , 19 , 20 ] ve elektronik sağlık kayıtlarının analizi [ 21 ] için veri madenciliğinde zamanla değişen verileri analiz etmede başarıyla kullanılmıştır . Ancak, dinamik metabolomik analizde tensör yöntemlerinin kullanımı, yakın zamana kadar bu tür uzunlamasına metabolomik verilerin eksikliği ve yöntemlerin metabolomikteki performansının sınırlı anlaşılması nedeniyle şimdiye kadar sınırlıydı. Bir istisna, sıçanlarda toksikolojik bir hasar üzerinde zaman içindeki tedavilerin etkisini incelemek için ASCA (ANOVA-eşzamanlı bileşen analizi) ile birleştirilmiş CANDECOMP/PARAFAC (CP) [ 22 , 23 ] tensör modelinin kullanılmasıdır [ 24 ].
Şekil 1
Bu makalede, dinamik metabolomik verileri analiz etmede ve altta yatan mekanizmaları ve dinamiklerini ortaya çıkarmada tensör faktörizasyonlarının potansiyelini araştırıyoruz. Bu tür yöntemlerin temel gerçeğini elde etmek ve sınırlamalarını ve avantajlarını incelemek için, yapılandırılmış doğrusal açık bir sistem, maya glikoliz modeli [ 25 ] ve insan kolesterol modeli [ 5 ] dahil olmak üzere artan karmaşıklığa sahip dinamik sistemlerin simülasyonları yoluyla veri üretiyoruz. Hem glikoliz modeli hem de kolesterol modeli in silico modeller. Bu in silico modeller, bir biyolojik sistemin gerçekçi modelleridir ve indüklenen varyasyonun farklı senaryolarını test etmeye olanak tanır. Gerçek verileri daha iyi taklit etmek için, denklemlerdeki kinetik parametreleri rastgele bozarak bu in silico modellere bireysel varyasyonu dahil ediyoruz ve ayrıca belirli parametrelerde bir azalma sağlayarak mutantları, yani indüklenen varyasyonu dahil ediyoruz. Simüle edilmiş verileri, Şekil 1’de gösterildiği gibi , özneler , metabolitler ve zaman modlarına sahip üç yönlü bir dizi olarak düzenliyoruz . Oluşturulan çok yönlü dizi daha sonra CANDECOMP/PARAFAC modeli olarak bilinen en popüler tensör modellerinden biri kullanılarak analiz edilir. CP modeli benzersiz olduğundan (permütasyon ve ölçekleme belirsizliklerine kadar) [ 13 , 27 ], diğer tensör modelleri yerine bu modeli seçtik, örneğin Tucker3 modeli [ 26 ]. Benzersizlik, dinamik metabolomik verileri analiz ederken önemli olan yorumlanabilir desenlere yol açar. Dahası, doğrusal olarak bağımlı faktörlerin varlığında gizli yapıyı CP modelinden daha iyi ortaya çıkarabildiğinden, kısıtlı bir CP modeli, yani Paralind (Doğrusal Bağımlılıklara Sahip Paralel Profiller) modelini [ 28 ] ele alıyoruz.
Yöntemler
Dinamik sistemler ve veri üretimi
Metabolit konsantrasyonlarının dinamikleri, aşağıdaki formdaki diferansiyel denklemlerle modellenebilir:
DXXDT=F(vv):=SSvv,XX(0)=XX0,(1)
Burada vektör metabolit konsantrasyonlarını temsil eder, türev metabolit konsantrasyonlarının zaman içindeki değişimini tanımlar, vektör metabolitler arasındaki reaksiyon akışlarını tanımlar ve matris metabolik ağı tanımlayan stokiyometrik matristir. Matristeki her satır bir metaboliti temsil eder, her sütun bir reaksiyona karşılık gelir ve her giriş, tüketilen metabolit ile negatif bir katsayı elde edilirken üretilen metabolit ile pozitif bir sayı verilecek bir reaksiyondaki bir metabolitin stokiyometrik katsayısını ifade eder. Vektör genellikle kinetik parametrelere sahip metabolitlerin konsantrasyonlarının bir fonksiyonudur.XXDXXDTvvSSSSvv
Doğrusal açık sistem
Akılar konsantrasyonların doğrusal fonksiyonları ise: , o zaman diferansiyel denklem şu şekilde yeniden yazılabilir: . 11 iç metabolit içeren doğrusal bir açık sistem oluşturuyoruz, burada ve boyutunda bir üç köşegen matristir . matrisindeki alt diyagonal elemanlar olarak ve süperdiyagonal elemanlar olması koşuluyla seçilir . Başlangıç değeri olarak ayarlanmıştır. Doğrusal açık sistem hakkında daha fazla ayrıntı Ek dosya 1’de bulunabilir : Bölüm 1. Verileri ürettiğimizde, simülasyonu [0, 0,2] dakikada ele alırız ve için zaman noktalarında çözümü seçeriz . Yol Ek dosya 1’de gösterilmiştir : Şekil S1.F(vv)=AAXX+BBDXXDT=AAXX+BBBB=103×[0,1,0,0,0,0,0,0,0,0,0,0]TAA11×11AA103×[0,2,0,1,0,5,0,3,2,1,3,0,4,1,0,4]T103×[0,3,0,5,2,2,0,3,3,0,5,1,0,2,0,4]TAA(11,11)=−103XX0=[1,1,1,1,1,1,1,1,1,1,1]T(6+5×k)×0,002k=0,1,…,19
Glikoliz modeli
Glikoliz modeli Van Heerden ve arkadaşları tarafından önerilmiştir [ 25 ] ve Denklem ( 1 )’deki doğrusal olmayan terim kinetik denklemleri tanımlayan parametreleri içerir. Bu model açık bir sistemdir ancak ek döngüler nedeniyle doğrusal açık sistemden çok daha karmaşıktır, örneğin metabolit FBP’den enzim PYK’ye ileri beslemeli kontrol döngüsü, ADP-ATP döngüsü ve Ek dosya 1’deki yol grafiğinde gösterilen NADH-NAD döngüsü : Şekil S3; bu model hakkında daha fazla ayrıntı Ek dosya 1’de bulunabilir : Bölüm 2. Verileri ürettiğimizde, [ 25 ]’te dikkate alınan varsayılan başlangıç değerlerini kullanırız. Simülasyonu [0, 0.2]dakikada ele alırızvvDipnot1içinzaman noktalarında çözümü seçin.(6+5×k)×0,002k=0,1,…,19
Kolesterol modeli
Kolesterol modeli van de Pas ve ark. tarafından önerildi [ 5 ] ve bu model için Denklem ( 1 )’deki doğrusal olmayan terim kinetik denklemlerdeki parametreleri içerir. Glikoliz modeline benzer şekilde, bu model de farklı kolesteroller arasında daha fazla döngüye sahip açık bir sistemdir; Ek dosya 1’deki yola bakın : Şekil S8. Model, örneğin ailevi hiperkolesterolemiye (FH), balık gözü hastalığına, Smith-Lemli-Opitz sendromuna (SLOS) ve diğer hastalıklara neden olan mutasyonlar dahil olmak üzere on bilinen mutasyona sahip verilerle doğrulandı [ 5 ]. Her mutasyon için, bazı belirli enzimler normal durumdan çok daha düşük aktivitelere sahiptir. Bu makalede, bu farklı mutant tiplerini indüklenen varyasyonların farklı kaynakları olarak ele alıyoruz. Verileri, [ 5 ] ‘tekiyle aynı başlangıç ayarlarını kullanarak modeli simüle ederek üretiyoruz , yani tüm normal denekler verilen başlangıç metabolit koşullarıyla başlıyor ve mutant denekler normal deneklerin sabit durum koşullarıyla başlıyor. Zaman noktalarını seçme şeklimiz şu şekildedir: [ 5 ]’te kullanılan zaman noktaları için, yani (logspace(0,6,1000)-1)vvDipnot2 , ilk zaman noktasından başlıyoruz ve toplamda 21 zaman noktası elde edene kadar her 24. zaman noktasını seçiyoruzDipnot3 .
Çok yönlü veri analizi
CANDECOMP/PARAFAC (CP) modeli
Bir tensörün poliadik formundan kaynaklanan CP modeli [ 30 ], 1970’te tanıtıldığından beri popüler hale gelmiştir [ 22 , 23 ]. CP çarpanlara ayırma, bir tensörü birinci rütbeli tensörlerin toplamı olarak gösterir (bkz. Şekil 1 ) ve matris Tekil Değer Ayrıştırması’nın (SVD) bir genellemesi olarak görülebilir. Üçüncü dereceden bir tensör in R bileşenli bir CP modeli aşağıdaki gibidir:XX∈RBEN×J×KXX
⟦⟧XX≈X^X^=⟦λλ;AA,BB,CC⟧:=∑R=1RλRAAR∘BBR∘CCR,
Burada birinci rütbe bileşenleri, sırasıyla faktör matrisleri olan ve vektörlerinden oluşmaktadır ve sütunlarıdır; bir skalerdir ve vektör dış çarpımını ifade eder. Bu tanımlamada, vektörü tarafından emildiği varsayılır . İki yönlü veri kümeleri için çoğu boyut indirgeme yönteminin aksine, CP modeli, ek kısıtlamalar getirmeden, hafif koşullar altında permütasyon ve ölçekleme belirsizliklerine kadar benzersizdir [ 13 , 27 ]. Benzersizlik, CP modelinin yorumlanabilir sonuçlar vermesini sağlar ve bu da onu yorumlanabilir veri analizi için çok tercih edilen bir araç haline getirir. Sonuçlar yorumlanırken, deneklerdeki faktör yüklemeleri , metabolitler ve zaman modları her bileşen için birlikte incelenmelidir.AAR,BBRCCRAA∈RBEN×R,BB∈RJ×RCC∈RK×RλR∘AA,BB,CCλλ
CP modeli , aşağıdaki optimizasyon problemini çözerek eksik girdileri olan verileri analiz etmek için de kullanılabilir [ 31 , 32 ]:
⟦⟧dakikaA,B,C‖BB∗(XX−⟦λλ;AA,BB,CC⟧)‖2,
Burada operatör Hadamard ürünüdür, girdileri aşağıdaki gibidir:∗‖.‖BB∈RBEN×J×K
En az bir modda doğrusal olarak bağımlı etkilere sahip varyasyonların temel kaynakları tarafından üretilen desenlere sahip üç yollu veriler için, en uygun CP modeli bu bağımlılıkları göstermelidir ve böyle bir çözüm rütbe eksiktir. Ancak, standart CP modeli verilerdeki gürültü nedeniyle gerçek gizli yapıyı ortaya çıkarmada başarısız olabilir [ 28 ]. Bunun yerine, başlangıçta kısıtlı bir Tucker modeli [ 33 ] olarak tanıtılan Paralind modeli [ 28 ] olarak adlandırılan özel bir CP modeli durumu daha uygundur. Bu model kısmen benzersizdir, yani yalnızca doğrusal olarak bağımsız faktör vektörlerine sahip faktörlerde benzersizliğe sahiptir ancak doğrusal olarak bağımlı faktörlerde benzersiz değildir. Verilerde bulunan örtük doğrusal bağımlılıkları açıkça temsil eder ve böylece gizli yapıyı daha doğru bir şekilde kurtarır. Ayrıca, Paralind modelinde daha az parametre kullanıldığından aşırı uyuma daha az eğilimlidir. İlk modda doğrusal olarak bağımlı faktörlere sahip Paralind modeli aşağıdaki gibi formüle edilebilir:
⟦⟧XX≈X^X^=⟦λλ;AA~,BB,CC⟧=∑R=1RλRA~R∘BBR∘CCR,
Burada ile ve , ve matrisi, doğrusal olarak bağımlı ilişkileri depolayan ‘bağımlılık matrisi’ olarak adlandırılır. Bu modeli Paralind( S , R , R ) ile gösteriyoruz . matrisi şu şekilde verilebilir:AA~=AAHHAA∈RBEN×SHH∈RS×RBB∈RJ×RCC∈RK×RHHHH
HH=[110001].
Sayısal deneyler
Bu bölümde, öncelikle veri setlerini oluşturmak için kullandığımız kurulumları sunacağız ve ardından CP ve Paralind modellerinin altta yatan mekanizmaları ve dinamikleri yakalama açısından performansını göstereceğiz.
Deneysel kurulum ve detaylar
Veri setlerini tanıtmadan önce, öncelikle bireysel ve indüklenen varyasyonları tanımlıyoruz.
Bireysel varyasyon, sabit kinetik parametrelere eklenen rastgele bozulmaları ifade eder. Bireysel varyasyonun seviyesi ( ile gösterilir ) bozulmaların seviyesine bağlıdır. Doğrusal sistem için, bireysel varyasyon, örneğin %1’lik bir seviye içinde süperdiyagonal ve subdiyagonal elemanlara rastgele bozulmalar eklenerek tanıtılır.βDipnotVarsayılan değerlerden 4’ü) ve her sütunun toplamlarının sıfır olarak tutulması, ancakher zaman uygulanırβ=0,01AA(11,11)=−1×103Dipnot5. Glikoliz ve kolesterol modelleri için, kinetik parametrelerin belirli bir seviyesi içinde, örneğin varsayılan değerlerin %2’si içinde () rastgele bozulmalar eklenerek bireysel varyasyon tanıtılır.β=0,02
İndüklenen değişim, belirli bir kinetik parametrede meydana gelen değişimi ifade eder ve indüklenen değişimin seviyesi ( ile gösterilir ) değişimin seviyesine bağlıdır.α
Aşağıdaki iki tür veri kümesini ele alacağız.
Bir kaynaklı indüklenen varyasyona sahip veri kümesi. Bu tür veri kümesi 20 denek içerir:
( Normal denekler) İlk 10 denek, yalnızca seviyesinde bireysel varyasyonla simülasyonların çalıştırılmasıyla elde edilmiştir ;β
( Anormal seviyesinde bireysel varyasyon değerinin %50 oranında azalmasını sağlayan indüklenen varyasyon ile simülasyonlar çalıştırılarak elde edilir (bu denekleri anormal_ A (7,6) denekleri olarak gösteriyoruz ); glikoliz modeli için, VmaxPFK’nin varsayılan değerlerinde %50 oranında azalma vardırβAA(7,6) Dipnot6 (bu konular abnormal_VmaxPFK konuları olarak gösterilir ); kolesterol modeli için, mutant1 (kullanılarak (bu konular abnormal_mutant1 ile gösterilirα=0,62Dipnot(7 konu).
İki kaynaklı indüklenen varyasyonlara sahip veri kümesi. Bu tür veri kümesi 30 denek içerir ve glikoliz ve kolesterol modelleri için üretilir:
( Normal denekler) İlk 10 denek, yukarıda açıklanan normal deneklerle aynı şekilde kullanılarak üretilir .β=0,02
( Anormal denekler) Bir sonraki 10 denek, glikoliz modelinde abnormal_VmaxPFK ( ) ve kolesterol modelinde abnormal_mutant6 ( dir .α=0,50α=0,35β=0,02
( Anormal denekler) Son 10 denek, glikoliz modelinde abnormal_VmaxPYK ( ) ve kolesterol modelinde abnormal_mutant10 ( değeri vardır .α=0,50α=0,95β=0,02
Her veri seti daha sonra özneler , metabolitler ve zaman modları ile üçüncü dereceden bir tensör olarak düzenlenir . Doğrusal açık sistem ve glikoliz modeli tarafından üretilen veri setleri, öznelerin sayısı × 11 metabolit × 20 zaman noktası boyutundadır ve kolesterol modeli tarafından üretilen veri setleri, öznelerin sayısı × 8 metabolit × 21 zaman noktası boyutundadır.
Veri ön işleme
Analizden önce, her üçüncü dereceden tensörü denek moduna göre merkezliyoruz [ 34 ]. Ayrıca, farklı metabolitlerin konsantrasyonları farklı aralıklarda olduğundan, tensör, metabolit modundaki her dilimin karekök ortalama değerine göre metabolit modunda ölçeklenir [ 34 ].
Model seçimi
Farklı modelleri değerlendirirken ve bileşen sayısını belirlerken, özellikle model uyumu, çekirdek tutarlılık tanısı, çapraz doğrulama ve Tucker’ın uyumluluk katsayısı olmak üzere çeşitli tanılamalar kullanırız. Model uyumu (genellikle açıklanan varyans olarak da adlandırılır ) şu şekilde tanımlanır:
Yerleştirmek=100×(1−‖XX−X^X^‖2‖XX‖2),
Burada ve sırasıyla orijinal verileri ve model tarafından veri yaklaşımını ifade eder. %100’lük bir uyum değeri, model tarafından tamamen açıklandığı anlamına gelirken, %100’den küçük bir uyum değeri, artıklarda açıklanamayan bir kısım kaldığı anlamına gelir. Farklı modeller (örneğin, farklı sayıda bileşene sahip modeller) için model uyumunda belirgin bir değişiklik, daha iyi bir model peşinde koşarken dikkate alınması gereken önemli bir kazanımı gösterir.XXX^X^XX
Çekirdek tutarlılık teşhisinin ayrıca bir CP modelindeki bileşen sayısını belirlemek için de yararlı olduğu gösterilmiştir [ 35 ]. Bir CP modelinin çekirdek tutarlılığı, çekirdek dizisinin süperdiyagonallik derecesinin karşılaştırılmasıyla tanımlanırDipnotCP modelinin 8’i ve CP faktörlerini kullanarak Tucker3 modeliyle [ 26 ] verilerin modellenmesiyle elde edilen çekirdek dizisi. %100’e yakın çekirdek tutarlılık değeri uygun bir modeli gösterir ve çok fazla bileşen kullanılırsa düşmesi beklenir.
Son olarak, model seçimi için çapraz doğrulama yoluyla eksik veri tahmin performansını kullanırız. Daha kesin olarak, verilere biraz gürültü ekleriz, yani,
XXgürültü=XX+ηNN‖XX‖‖NN‖,
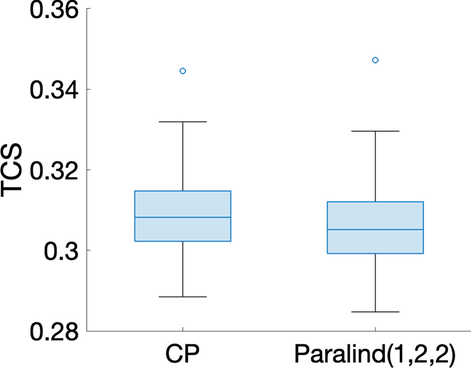
burada girişleri standart normal dağılımdan rastgele çekilen üçüncü dereceden bir tensördür ve gürültü seviyesidir. Tensör girişlerinin %20’sini rastgele eksik olarak ayarladık, verileri önceden işledik ve eksik girişleri kurtarmak için farklı modeller (yani, CP ve Paralind) kullandık. Farklı rastgele eksik giriş kümelerini kullanarak yöntemlerin performansını değerlendirmek için bu işlemi 20 kez tekrarladık. Daha sonra farklı modellerin performansı, [ 31 ] olarak tanımlanan tensör tamamlama puanı (TCS) kullanılarak değerlendirildiNNη
TÇS=‖(1−BB)∗(XX^−XXgürültü)‖‖(1−BB)∗XXgürültü‖,
Burada Denklem ( 2 ) ile tanımlanır . TCS, bir model için test hatasının bir değerlendirmesi olarak görülebilir ve daha düşük bir değer, modelin verilerdeki temel kalıpları yakalamada daha iyi davrandığını gösterir.BB
CP modelleri iki faktörlü bir dejenerasyondan muzdarip olabilir (dejenerasyon hakkında daha fazla ayrıntı için [ 36 ]’ya bakın). Modelin iki faktörlü bir dejenerasyona sahip olup olmadığını değerlendirmek için Tucker’ın uyumluluk katsayısını (TC ile gösterilir) kullanırız [ 37 ]. i’inci ve j’inci bileşen için TC değeri şu şekilde tanımlanır:
Bu, her moddaki iki bileşenin kosinüs benzerliğinin ( ) çarpımına karşılık gelir. Bu makalede, TC değerini olarak alıyoruz; burada . – 1’e yakın bir TC değeri, geçerli bir model olmayan dejeneratif bir modeli gösterir.CBenJ=AABenTAAJ‖AABen‖‖AAJ‖TC=TCBen0J0|TCBen0J0|=maksimumBen,J|TCBenJ|
Uygulama detayları
CP modelleri , Tensor Toolbox sürüm 3.1’den [ 39 ] cp-opt [ 38 ] ve cp-wopt [ 31 ] (eksik girişleri olan verilere) kullanılarak , sınırlı bellekli BFGS (LBFGS-B) kullanılarak uygulanırDipnot9 optimizasyon algoritması olarak. Zaman modunda negatif olmayan kısıtlamayı uygularız. Paralind modeliDipnot10, Bro ve diğerleri tarafından tanıtılan algoritma kullanılarak yerleştirilir [ 28 ]. Benzersiz modeller elde etmek için, Paralind modelini yerleştirirken metabolitler modunda faktör matrisinin ortogonal ve zaman modunda negatif olmayan olmasını zorunlu kılıyoruz. Yerel minimumlardan kaçınmak için çoklu rastgele başlatmalar kullanılır. Çekirdek tutarlılığının hesaplanması için, N-yollu araç kutusundan corcond fonksiyonunu kullanırız [ 40 ]. Tüm deneyler MATLAB’da (2020a sürümü) gerçekleştirilir.
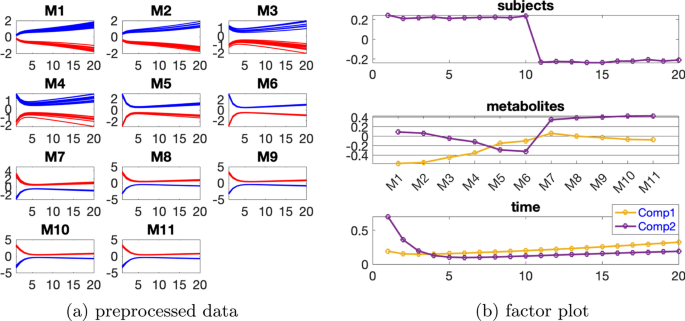
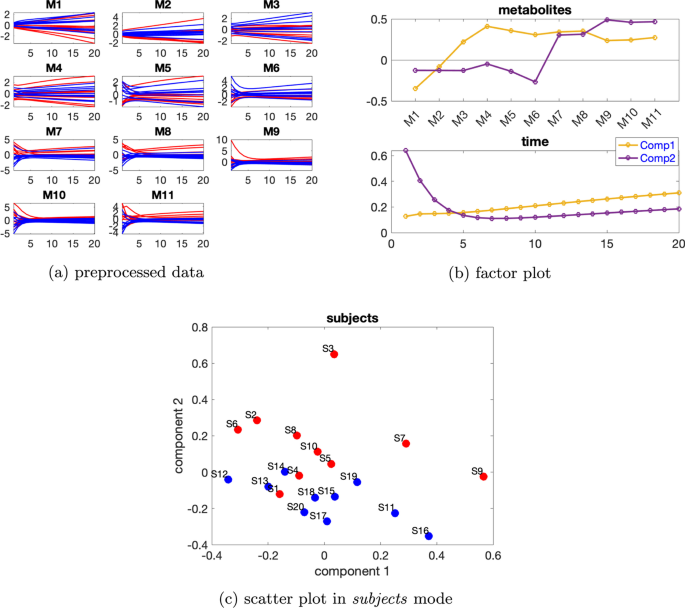
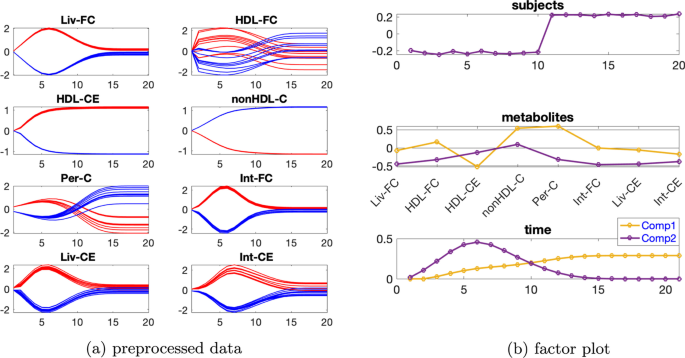
Paralind(1,2,2) modeli verilerin %98,38’ini açıklıyor ve bu, ekstra kısıtlama nedeniyle CP modelinden biraz daha düşük. Paralind(1,2,2) modelinin özne modu, iki grup arasında net bir ayrım gösteriyor (Şekil 3 b). Metabolitler modundaki ilk bileşenden (Şekil 3 b), M1 , M2 , M3 ve M4 metabolitlerinin büyük mutlak katsayılara sahip olduğunu ve zaman modunda ilk bileşenin bu metabolitlerde gösterilen dinamikleri yakaladığını gözlemliyoruz . Bu bileşen için kalan metabolitlerin katsayıları sıfıra yakındır. İkinci bileşen için, tam tersi şekilde, M7 , M8 , M9 , M10 , M11 ve M5 , M6 metabolitleri büyük katsayılara sahiptir ve bu metabolitlerde gösterilen dinamikler, zaman modunda ikinci bileşen tarafından yakalanır. Ayrıca, metabolitler modundaki her iki bileşenden (Şekil 3 b), Şekil 3 a’da gösterilen bu iki metabolit arasındaki mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan, metabolitler M6 ve M7 arasında bir sıçrama değişimi gözlemliyoruz . Bu değişim, anormal_ A (7,6) deneklerinde nın azalmasından kaynaklanmaktadır ve değişimin model tarafından başarılı bir şekilde yakalanması, Şekil 3 b’de görüldüğü gibi, denek modunda normal (ilk 10 denek) ve anormal_ A (7,6) (son 10 denek) gruplarının başarılı bir şekilde ayrılmasıyla sonuçlanır .AA(7,6)
Şekil 3
Daha büyük bireysel varyasyon dikkate alındığında, rütbe eksikliği ortadan kalkar ve CP modelleri altta yatan örüntüleri daha iyi yakalar, örneğin bkz. Şekil 4 , burada bireysel varyasyon seviyesi ve 2 bileşenli CP modeli için denek modundaki iki bileşenin benzerlik puanı ‘tür . CP modeli verilerin %62,42’sini açıklar. Şekil 4b’de sunulan metabolitler ve zaman modlarındaki ilk bileşenden, M2 hariç tüm metabolitlerin büyük mutlak değerlere sahip katsayılara sahip olduğunu ve zaman modundaki bileşenin tüm metabolitlerde görülen dinamiği bir dereceye kadar yakaladığını gözlemliyoruz. Metabolitler modundaki ikinci bileşenden ( 4b ), M6 , M7 , M8 , M9 , M10 ve M11 metabolitlerinin büyük katsayılara sahip olduğunu ve zaman modunda bu bileşenin bu metabolitlerde gösterilen hızlı düşüşü yakaladığını gözlemliyoruz . M9 , M10 ve M11’deki dinamikler esas olarak ikinci bileşen tarafından yakalanır, ancak metabolitler M5 , M6 , M7 ve M8’deki dinamikler, Şekil 4a’da gösterildiği gibi, zaman modunda iki bileşenin bir karışımıdır . Ayrıca, Şekil 3b’de gösterilen sıçrama değişimine benzer şekilde, metabolitler modunda ikinci bileşende metabolitler M6 ve M7 arasında bir sıçrama değişimi gözlemliyoruz ( 4b ). Bu , Şekil 4a’da gösterilen metabolitler M6 ve M7’deki mavi ve kırmızı çizgilerin değişimiyle tutarlıdır ve anormal_ A (7,6) deneklerinde ‘nın azalmasından kaynaklanmaktadır . Bu nedenle, denek modundaki ikinci bileşenin, Şekil 4c’de gösterildiği gibi, normal ve anormal_ A (7,6) denekleri bir dereceye kadar ayırabilmesi makuldür.β=0,3C12=−0,24AA(7,6)
Şekil 4
Yardımcı deneyler, CP modellerinin davranışının kinetik katsayılara dayandığını göstermektedir. Bazı özel durumlar için, CP modelleri için dejenerasyon gözlemlenmiştir, örneğin, ayarı ve köşegen elemanlarının 1 , 1, 1, 1, 1, 1]\) olarak ayarlandığı bir üç köşegen matris \({\varvec{{A}}} . Böyle bir ayar ve küçük bir bireysel varyasyonla doğrusal açık sistem tarafından üretilen veriler için, örneğin, , CP modeli dejeneratiftir. Ancak, Paralind modeli bu gibi durumlarda da yararlıdır ve altta yatan dinamikleri yakalar; bkz. Ek dosya 1 : Şekil S2.BB=103×[0,5,0,0,0,0,0,0,0,0,0,0]TAA103×[−1,−2,−2,−2,−2,−2,−2,−2,−2,−2,−2]103×[1,1,1,1,1,1,1,1,1,1]β=0,01
Glikoliz modeli
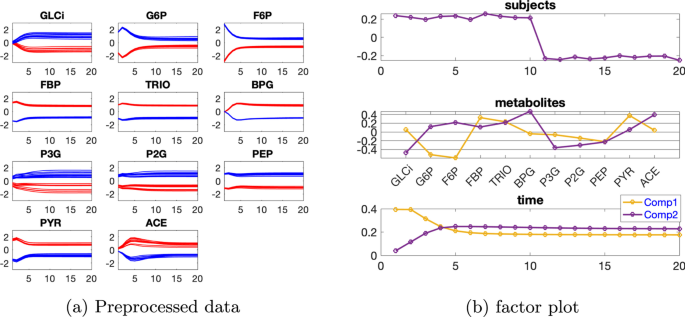
Tek kaynaklı indüklenen varyasyona sahip veri seti
Veriyi, VmaxPFK’nin varsayılan değerinin %50 azalması ve bireysel varyasyonu seviyesindeki indüklenmiş varyasyon olarak ele alıyoruz. Her bir metabolitin zamansal profilleri Şekil 5a’da gösterilmiştir . Tablo 2’ye dayanarak , 2 bileşenli bir CP modeli kullanıyoruz ve doğrusal açık sistemde olduğu gibi, denek modunda rütbe eksikliği gözlemliyoruz . Rütbe eksikliğini hesaba katmak için, bu veri setini analiz etmek için bunun yerine bir Paralind(1,2,2) modeli kullanıyoruz. CP’nin Paralind’e karşı çapraz doğrulama performansı, verilerin %96,05’ini açıklayan Paralind(1,2,2) modelinin bu veri seti için daha iyi bir seçim olduğunu da gösteriyor (bkz. Ek dosya 1 : Şekil S4). İki denek grubu iyi ayrılabilir ve doğrusal sistemle karşılaştırıldığında, Şekil 5b’de gösterilen metabolit modundaki faktör grafiği, ağın karmaşıklığı nedeniyle daha karmaşıktır. β=0,02Tablo 2 Açıklanan varyans (uyum), çekirdek tutarlılık (CC), Tucker’ın uyumluluk katsayısı (TC), ilk iki bileşenin kosinüs benzerlik puanı ( ) denek modunda ve bileşen sayısı ( R ) glikoliz modeli tarafından üretilen verileri analiz etmek için kullanılan CP modelleri için tek kaynaklı indüklenmiş varyasyon ve bireysel varyasyon düzeyindeC12β=0,02
İlk bileşen, Şekil 5a’da gösterilen bu metabolitler arasındaki mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan ve VmaxPFK’nin azalmasından kaynaklanan, biri F6P ve FBP metabolitleri arasında, diğeri PEP ve PYR metabolitleri arasında olmak üzere iki büyük sıçrama değişikliği olduğunu göstermektedir . F6P ve FBP metabolitleri arasındaki değişiklik, Şekil 3’te doğrusal açık sistem için gösterilen değişikliğe benzer şekilde , doğrudan VmaxPFK’nin azalmasına karşılık gelir. PEP ve PYR metabolitleri arasındaki değişiklik, VmaxPFK’nin azalması ve yol grafiğinde gösterilen ileri beslemeli kontrol döngüsünün neden olduğu FBP azalmasından kaynaklanan VmaxPYK enziminin aktivitesinin azalmasına karşılık gelir (Ek dosya 1 : Şekil S3). Metabolitler G6P , F6P , FBP ve PYR ilk bileşende büyük mutlak katsayılara sahiptir ve bu metabolitlerde gösterilen üçüncü zaman noktalarından gelen dinamikler, Şekil 5b’de gösterildiği gibi, zaman modunda ilk bileşen tarafından iyi yakalanmıştır. Metabolitler FBP ve PYR’de gösterilen çıkıntılar, zaman modunda iki bileşenin doğrusal kombinasyonları tarafından yakalanabilir. Ek bir bileşene sahip modeller, örneğin Ek dosya 1’de gösterildiği gibi G6P’deki çıkıntı gibi daha fazla varyansı yakalamak için yararlı olacaktır : Şekil S5. Ancak, dinamik varyasyonların çoğunu yakaladığı ve yorumlanması daha kolay olduğu için Paralind (1,2,2) modelini kullanmayı tercih ediyoruz.
Metabolitler modundaki ikinci bileşen, Şekil 5a’da gösterilen mavi ve kırmızı çizgilerin geçişiyle tutarlı olan metabolitler BPG ve P3G arasında bir sıçrama değişimini gösterir. Bu geçiş, VmaxPYK reaksiyon hızının düşmesiyle oluşan PEP , P2G ve P3G artışından kaynaklanır.Dipnot11 ve anormal_VmaxPFK denekleri için VmaxPFK’nin azalması nedeniyle FBP , TRIO ve BPG’nin azalması. Metabolitler GLCi , BPG ve ACE ikinci bileşende büyük mutlak puanlara sahiptir ve bu metabolitlerin dinamikleri, Şekil 5b’de gösterildiği gibi zaman modunda ikinci bileşen tarafından iyi yakalanmıştır . Metabolitler TRIO , P3G , P2G ve PEP’de gösterilen dinamikler , zaman modunda her iki bileşenin bir karışımıdır.
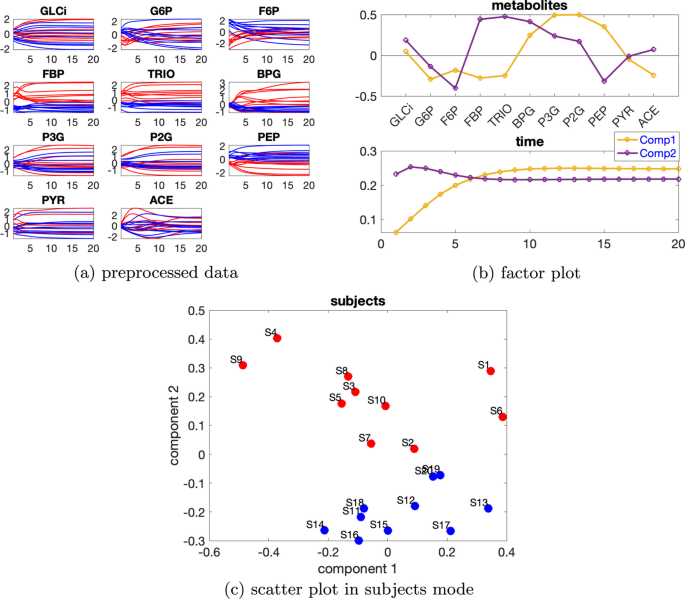
Daha yüksek düzeyde bireysel varyasyon düşünüldüğünde, özneler modundaki doğrusal bağımlılık zayıflar, örneğin bkz. Ek dosya 1 : Tablo S1, burada bireysel varyasyon düzeyi ve özneler modundaki iki bileşenin kosinüs benzerlik puanı 2 bileşenli bir CP modeli için dir . Bu nedenle Paralind modelleri yerine CP modelleri tercih edilir. Ek dosya 1 : Tablo S1’den, çekirdek tutarlılık değerleri 2 veya 3 bileşenli bir modelin kullanıldığını göstermektedir. 3 bileşenli modeldeki ek faktör yararlı bilgi sağlamadığından 2 bileşenli CP modelini seçtik. 2 bileşenli CP modeli verilerin %54,12’sini açıklıyor. Metabolitler ve zaman modundaki ilk bileşenden (bkz. Şekil 6b ), P3G , P2G ve PEP metabolitlerinin büyük katsayılara sahip olduğunu ve Şekil 6a’da gösterildiği gibi bu metabolitlerin dinamiklerinin yakalandığını gözlemliyoruz. Metabolitler modundaki ikinci bileşenden (bkz. Şekil 6b ), F6P , FBP , TRIO ve BPG metabolitlerinin büyük mutlak katsayılara sahip olduğunu ve bu metabolitlerin çoğunda mavi ve kırmızı çizgilerin ayrılabilir olduğunu gözlemliyoruz . Bu, Şekil 6c’de gösterildiği gibi, normal ve abnormal_VmaxPFK denekleri arasında ikinci bileşen tarafından denek modunda gözlemlenen ayrımla tutarlıdır. Zaman modundaki ikinci bileşen, bu metabolitlerdeki bazı deneklerin gösterdiği dinamikleri yakalar (Şekil 6a ).β=0,36C12=−0,28
Şekil 6
Daha da büyük bireysel varyasyona sahip veriler için, CP modelleri grupları ayıramayabilir. Başarısızlık, (i) bireysel varyasyonun varyansa hakim olmasından kaynaklanır, örneğin Ek dosya 1’e bakın : Şekil S6, burada bireysel varyasyon seviyesi, indüklenen varyasyona eşittir ( ), (ii) bir veya iki deneğin kendine özgü davranış gösterme olasılığı olan sınırlı sayıda denek ( Ek dosya 1’deki BPG profillerine bakın : Şekil S6a) ve böylece ortaklık çıkarmak zorlaşır. Gerçekten de, denek sayısı daha fazla olduğunda (Ek dosya 1’e bakın : Şekil S7), verilerin %66,56’sını açıklayan 3 bileşenli CP modeli, verilerdeki ana değişimi ( VmaxPFK’nin azalması) yakalayabilir ve normal ve anormal_VmaxPFK için bile başarıyla ayırabilir . İdiopatik davranışların daha yaygın hale gelmesi, modellemeyi kolaylaştırmıştır.β=α=0,50β=α=0,50
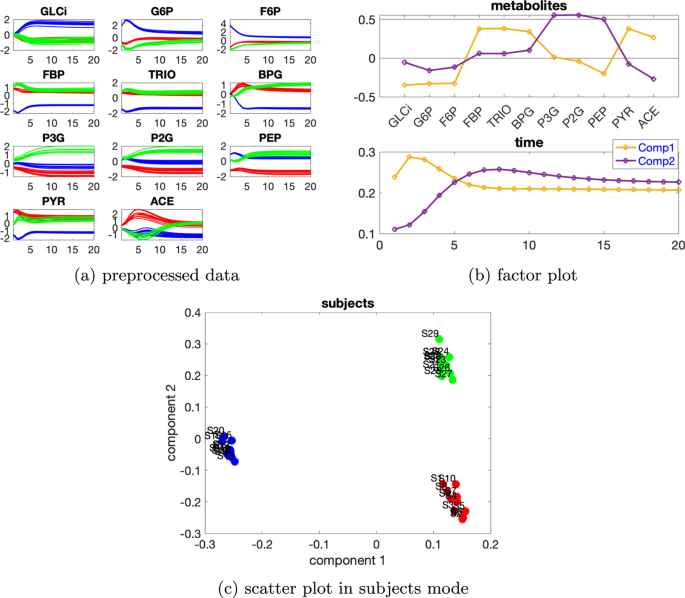
İki kaynaklı indüklenen varyasyona sahip veri setiseviyesindeki bireysel varyasyonve iki kaynaklı indüklenen varyasyon ile üretilen verileri VmaxPFK için varsayılan değerin %50 azalması ve VmaxPYK için varsayılan değerin %50 azalması. Tablo 3, verilerin %88,68’ini açıklayan 2 bileşenli bir CP modelinin kullanıldığını göstermektedir.β=0,02Tablo 3 Açıklanan varyans (uyum), çekirdek tutarlılık (CC), Tucker’ın uyumluluk katsayısı (TC), ilk iki bileşenin kosinüs benzerlik puanı ( ) denek modunda ve bileşen sayısı ( R ) glikoliz modeli tarafından üretilen verileri analiz etmek için kullanılan CP modelleri için VmaxPFK’de %50 azalma ve VmaxPYK’de %50 azalma olarak iki kaynaklı indüklenmiş varyasyon ve ayrıcaC12β=0,02
Metabolitlerdeki ve zaman modundaki ilk bileşenden (bkz. Şekil 7b ), GLCi , G6P , F6P , FBP , TRIO , BPG , PYR ve ACE metabolitlerinin büyük mutlak katsayılara sahip olduğunu ve Şekil 7a’da gösterilen bu metabolitlerin çoğundaki dinamiklerin yakalandığını gözlemliyoruz. Ayrıca, Şekil 7a’da gösterildiği gibi, mavi çizgiler bu metabolitlerdeki diğer çizgilerden ayrılabilir . Bu, denek modundaki ilk bileşenin (Şekil 7c ) abnormal_VmaxPFK deneklerini diğerlerinden ayırdığı gözlemiyle tutarlıdır. Dahası, Şekil 7a’daki VmaxPFK azalması ve ileri beslemeli kontrol döngüsü nedeniyle mavi çizgilerin diğer çizgilerle yer değiştirmesine uygun olarak, F6P ve FBP metabolitleri ile PEP ve PYR arasında bir sıçrama değişimi gözlemliyoruz . Bu gözlemler, Şekil 5 a, b’de glikoliz modeli için bir kaynaklı indüklenen varyasyon için fark edilenlere benzerdir . Metabolitler ve özneler modundaki ikinci bileşenden (bkz. Şekil 7 b, c), P3G , P2G ve PEP metabolitlerinin büyük puanlara sahip olduğunu ve üç tür öznelerin birbirinden ayrılabileceğini görüyoruz . Bu, Şekil 7 a’da gösterildiği gibi , P3G ve P2G metabolitlerinde farklı renklerde çizgilerin ayrılabilmesi nedeniyle mantıklıdır. Dahası, metabolitler modunda bu bileşende PEP ve PYR metabolitleri arasında bir sıçrama değişimi gözlemliyoruz . Bu, Şekil 7 a’da gösterilen diğer çizgilerle yeşil çizgilerin yer değiştirmesiyle uyumludur ve VmaxPYK’nin azalmasından kaynaklanmaktadır . Zaman modunda, P3G ve P2G metabolitlerinde gösterilen dinamiklerin ikinci bileşen tarafından yakalandığını gözlemliyoruz .
Kolesterol modeli
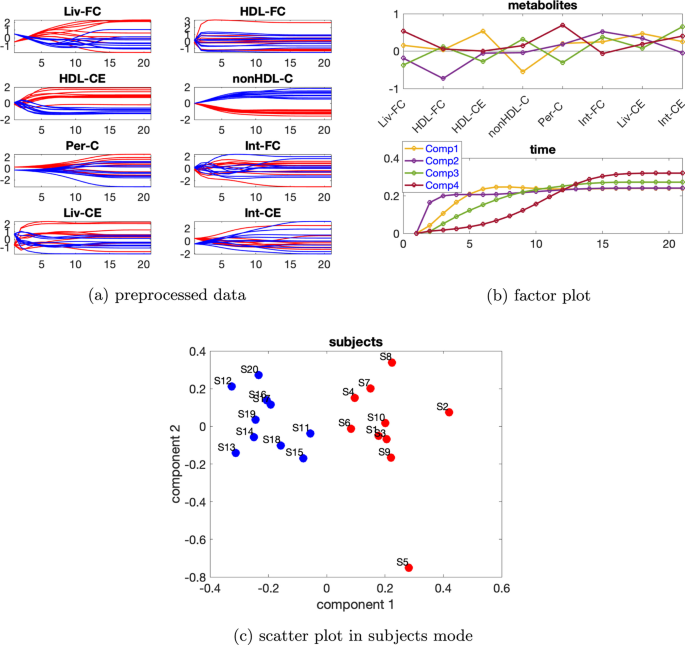
Tek kaynaklı indüklenmiş varyasyona sahip veri seti İndüklenmiş varyasyona sahip verileri mutant1 ve bireysel varyasyonu. Önceden işlenmiş verilerin zamansal profilleri Şekil 8a’da gösterilmiştir . Ek dosya 1 : Tablo S2, 2 veya 3 bileşenli bir modelin kullanıldığını gösterir ve hem iki hem de üç bileşene sahip CP modelleri için denek modunda rütbe eksikliği gözlemlenirβ=0,02
Şekil 8
Bu nedenle Paralind modelini kullanıyoruz ve yorumdan, 2 bileşenli bir modeli tercih ediyoruz. Dahası, çapraz doğrulama performansı (Ek dosya 1 : Şekil S9), Paralind(1,2,2) modelinin 2 bileşenli CP modelinden daha iyi davrandığını göstermektedir. Paralind(1,2,2) modeli verilerin %89,10’unu açıklamaktadır. Özneler modundaki faktör grafiğinden ( Şekil 8 b), normal ve abnormal_mutant1 özneleri arasında net bir ayrım görüyoruz . Metabolitler ve zaman modundaki ilk bileşenden (bkz. Şekil 8 b), HDL-CE , nonHDL-C ve Per-C metabolitlerinin büyük katsayılara sahip olduğunu, kalan metabolitlerin katsayılarının ise sıfıra yakın olduğunu gözlemliyoruz ; zaman modundaki bileşen, metabolit nonHDL-C’de gösterilen dinamikleri ve ayrıca Şekil 8 a’da gösterildiği gibi HDL-CE ve Per-C metabolitlerindeki dinamiklerin bir karışımını yakalar . Ayrıca, Şekil 8a’daki mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan metabolitler HDL-CE ve nonHDL-C arasında net bir sıçrama değişimi gözlemliyoruz ve bu, mutant1’in neden olduğu abnormal_mutant1 denekleri için metabolitler nonHDL-C’nin yükselmesi ve metabolitler HDL-CE’nin azalmasından kaynaklanmaktadır. Metabolitler ve zaman modundaki ikinci bileşenden (bkz. Şekil 8b ), Liv-FC , Int-FC , Liv-CE ve Int-CE metabolitlerinin büyük katsayılara sahip olduğunu ve zaman modundaki bileşenin , Şekil 8a’da gösterildiği gibi, bu metabolitlerde gösterilen ortak dinamikleri yakaladığını görüyoruz ; ayrıca, Şekil 8a’da gösterilen mavi ve kırmızı çizgilerin yer değiştirmesiyle tutarlı olan metabolitler nonHDL-C ve Per-C arasında bir sıçrama değişimi gözlemliyoruz .
Şekil 9
Bu değişim aynı zamanda mutant1’den de kaynaklanmaktadır çünkü nonHDL-C’den Liv-FC ve Per-C’ye olan reaksiyon hızı azalır ve bu da nonHDL-C’nin artmasına ve Per-C’nin azalmasına yol açar .
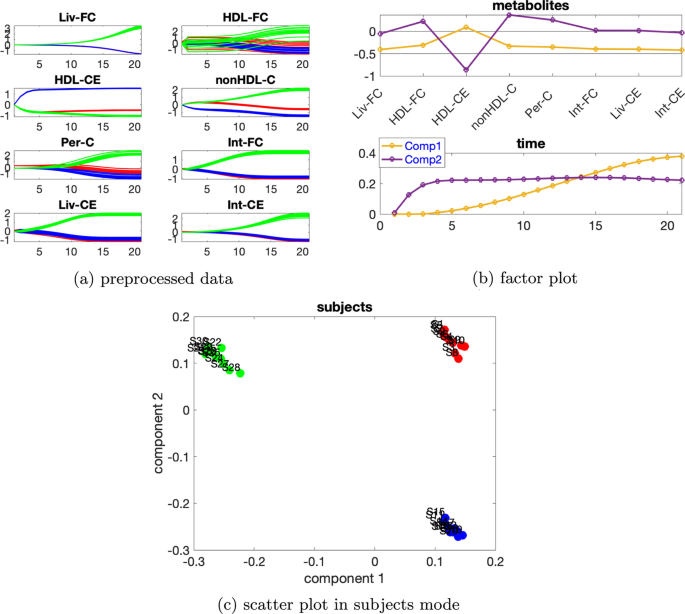
Yüksek düzeyde bireysel varyasyonlar dikkate alındığında, denekler modundaki rütbe eksikliği ortadan kalkar ve CP modelleri tercih edilir. düzeyinde bireysel varyasyona sahip verileri ele alıyoruz. Ek dosya 1 : Tablo S3’e dayanarak , verilerin %79,15’ini açıklayan 4 bileşenli bir CP modeli kullanıyoruz. Metabolitlerdeki ve zaman modundaki ilk bileşenden (bkz. Şekil 9 a), HDL-CE ve nonHDL-C metabolitlerinin en büyük mutlak katsayılara sahip olduğunu ve Şekil 9 b’de gösterildiği gibi metabolit HDL-CE’deki dinamiklerin yakalandığını gözlemliyoruz . Ek olarak, Şekil 9 c’de denekler modundaki ilk bileşenin normal ve abnormal_mutant1 denekleri ayırdığını görüyoruz . Bu mantıklıdır çünkü mavi ve kırmızı çizgiler Şekil 9 a’da gösterildiği gibi metabolitler HDL-CE ve nonHDL-C’de ayrılabilir. Zaman modundaki ikinci bileşen, metabolitler modunda ikinci bileşende en önemli mutlak puanı alan metabolit HDL-FC’de gösterilen dinamikleri yakalar . Üçüncü bileşen, metabolitler modunda üçüncü bileşende en büyük pozitif puanı alan metabolit Int-CE’de gösterilen dinamikleri yakalar ve dördüncü bileşen , metabolitler modunda dördüncü bileşende en büyük pozitif puanı alan metabolit Per-C’de gösterilen dinamikleri yakalar .β=0,65
Şekil 10
İki kaynaklı indüklenmiş varyasyona sahip veri kümesi Bireysel varyasyonla[ 5 ]’teki mutant6 ve mutant10 olarak iki kaynaklı indüklenmiş varyasyonu. Ek dosya 1 : Tablo S4, 2 bileşenli bir modelin kullanıldığını gösteriyor. İki bileşenli CP modeli verilerin %91,89’unu açıklıyor.Şekil 10c’de gösterilen denek modundan , ilk bileşenin abnormal_mutant10 deneklerini kalan deneklerdenayırdığını , ikinci bileşenin ise normal denekleri abnormal_mutant6 deneklerinden ayırdığını gözlemliyoruz. Bu mantıklıdır çünkü HDL-CE hariç tüm metabolitlerin metabolit modunda ilk bileşen üzerinde büyük bir katsayıları vardır ve Şekil 10a’dan bu metabolitler için önceden işlenmiş verilerdeki mavi çizgilerin ve kırmızı çizgilerin oldukça yakın olduğunu ve yeşil çizgilerden açıkça ayrıldığınıgörebiliriz Metabolit HDL-CE ikinci bileşende en büyük mutlak puana sahipken ve mavi çizgiler,Şekil 10 a’da gösterildiği gibi metabolit HDL-CE için diğer çizgilerden açıkça ayrılabilir. Metabolitlerdeki ve zaman modundaki grafikleri birleştirdiğimizde (Şekil 10 b), modelin iki ana tür dinamiği yakaladığını gözlemliyoruz, yani metabolit HDL-CE’de gösterilen sabit duruma (ikinci bileşen) hızla artanve kalan metabolitlerin çoğunda gösterilen sabit duruma (birinci bileşen) doğru yavaşça artan.β=0,02
Çözüm
Bu makalede, dinamik sistemlerin simülasyonları yoluyla üretilen dinamik metabolomik verilerinin analizi için tensör faktörizasyonlarını inceledik. CP ve Paralind modeli de dahil olmak üzere bu tür yöntemlerin temel fikri, konular arasındaki ortaklığı, yani ortak dinamik davranışları çıkarmaktır. Pratikte karşılaşılan metabolik sistemlerin dinamik davranışı (i) farklı varyasyon kaynaklarının boyutlarına ve (ii) sistemin kendisinin yapısına, yani metabolik ağın topolojisinin yanı sıra kinetik sabitlerin boyutlarına bağlıdır. Artan karmaşıklığa sahip dinamik sistemleri, yani doğrusal açık bir sistem, bir maya glikoliz modeli ve bir insan kolesterol modeli kullanarak, sistemin yapısını ve farklı varyasyon kaynaklarını inceledik ve CP ve Paralind modellerinin farklı ortamlarda altta yatan dinamikleri ne kadar iyi yakaladığını gösterdik. İncelediğimiz yeterli ortaklığın olduğu tüm durumlarda, üç yollu verileri nispeten basit çok yollu modellerle, yani CP ve Paralind modelleriyle modelleyebiliriz. Bu modeller, metabolitler arasındaki ilişkilerdeki değişikliklerin başarılı bir şekilde yakalanmasıyla yansıtılan verilerdeki müdahaleleri tespit etmeyi başarır; bu, metabolitlerin faktör grafiklerindeki sıçrama değişiklikleriyle gösterilir. Metabolik ağ (topoloji ve bağlantı güçleri) ile CP veya Paralind modellerindeki metabolitlerin faktör yüklemeleri arasındaki ilişkinin ayrıntılı bir açıklaması, takip araştırmasının konusudur. Çoğu durumda, altta yatan silico modelden çıkarılan kalıpları da açıklayabilir ve anlayabiliriz. Ancak, dinamik davranıştaki bireysel farklılıklar pratikte, örneğin meydan okuma testlerinde muazzam olabilir [ 41 ]. Bu, örneklenen sınırlı sayıda bireyde, kendine özgü davranışa sahip bazılarının olacağı anlamına gelir. Deneylerimizde, bu kendine özgü davranışın daha çok bir yetersiz örnekleme sorunu olduğunu gösterdik.
CP ve Paralind modeli arasındaki seçim veri özelliklerine bağlıdır ve bu da, yukarıdaki paragrafta tartışılan (i) ve (ii) iki yönüne bağlıdır. Bu makalede, pratikte uygun bir model seçmek için iyi tanılamalar sunuyoruz. Küçük bireysel varyasyona sahip veriler ve dinamik davranış üzerinde benzer etkilere sahip indüklenen varyasyon kaynakları için Paralind modelini kullanıyoruz (CP modelindeki doğrusal bağımlılık faktörleri nedeniyle); büyük bireysel varyasyona sahip veriler veya çeşitli indüklenen varyasyonlara sahip veriler için CP modellerinin iyi çalıştığını gösteriyoruz.
Gecikmeli dinamik sistemler veya indüklenen varyasyondaki önemli farklılıklardan dolayı farklı dinamiklere sahip sistemler veya büyük kendine özgü davranışlar gibi daha karmaşık durumlar için, PARAFAC2 [ 42 ] veya Sınırlı Tucker [ 33 ] gibi daha karmaşık çok yönlü modellere ihtiyaç duyabiliriz. Ayrıca, zamanla evrimleşen metabolitlerle ilgilendiğimiz durumlar için [ 10 ], PARAFAC2’nin metabolitler modunda evrimleşen faktör matrislerini yakalayarak bunları ortaya çıkarması beklenir . Ayrıca, bireyler arasındaki rastgele varyasyonu hesaba katan karışık etkili üç yönlü modelleri dikkate almaya değer olabilir.
Bu simülasyon çalışması gerçek bir dinamik metabolomik veri setinin analizinden ilham almıştır. Gerçek verilerde, altta yatan dinamik ağ bilinmemektedir ve veri seti boyutu daha büyüktür, örneğin, metabolit ve denek sayısı yüzlerce mertebesindedir. CP modellerinin hala varyasyonların ana modellerini ve karşılık gelen zamansal profilleri ortaya çıkarması beklenmektedir, bunu gerçek bir metabolomik meydan okuma testi veri setindeki bulgularımızla göstermeyi planlıyoruz. Gelecekte bu tür büyük ölçekli dinamik metabolomik veri setleri mevcut olacaksa, yöntemler daha büyük veri setlerine [ 43 , 44 ] (her modda binlerce veya daha fazla değişkenle) de ölçeklenebilir.
Glikoliz modelindeki reaksiyonlar çok hızlıdır ve metabolitlerin konsantrasyonları sabit duruma hızla ulaşır. Bu nedenle, dinamik değişimin ortaya çıktığı kısa bir zaman aralığına odaklanıyoruz. Ancak, böyle bir zaman ölçeğinde gerçek metabolomik verileri edinmek mümkündür; hücre içi metabolitleri çıkarırken örnekleme süresinin örnek başına 220 ms olabileceği [ 29 ]’a bakın.
MATLAB gösteriminde: tspan=logspace(0,6,1000)-1 vektörünü ele alalım; seçilen zaman noktaları tspan(1:24:500)’dir.
Küçük biyolojik varyasyonlara sahip sistemleri taklit etmek için küçük sayılar seçiyoruz, ancak bu sayı, makalede daha sonra tartışılacağı üzere büyük de olabilir.
Bu kısıtlamalar doğrusal açık sistemlerde kütle korunum yasasının sağlanması için veri üretiminde her zaman kullanılır.
Burada, VmaxPFK dışındaki enzimler de düşünülebilir. Yolun orta kısmında konumlanmış bir enzimle başlamak istediğimiz için VmaxPFK’yi seçiyoruz.
İndüklenen varyasyon diğer enzim reaksiyon hızlarındaki azalmayla, örneğin VmaxPYK / mutant6 / mutant10’un %50 azalmasıyla tanımlanıyorsa , anormal denekler sırasıyla abnormal_VmaxPYK / abnormal_mutant6 / abnormal_mutant10 olarak gösterilir .
Bir CP modelinin çekirdek dizisi, CP modelini Tucker3 modelinin özel bir durumu olarak ifade ederek elde edilen çekirdek tensörüdür. CP modelinin çekirdek dizisi, , yani, rütbe bir bileşenlerinin ağırlıkları, süperdiyagonalde ve diğer tüm girişler sıfır olan bir süperdiyagonal tensördür.λ
VmaxPYK reaksiyon hızının azalması, VmaxPFK’nin azalmasından ve bunun sonucunda FBP’nin azalmasından ve ileri beslemeli kontrol döngüsünün etkisinden kaynaklanmaktadır.
Referanslar
Pellis L, van Erk MJ, van Ommen B, Bakker GC, Hendriks HF, Cnubben NH, Kleemann R, van Someren EP, Bobeldijk I, Rubingh CM, ve diğerleri. Yemek sonrası bir meydan okumadan sonra plazma metabolomikleri ve proteomik profillemesi, insan metabolik durumu üzerinde ince diyet etkilerini ortaya koymaktadır. Metabolomik. 2012;8(2):347–59.MaddeCASGoogle Akademik
van Duynhoven J, Vaughan EE, Jacobs DM, Kemperman RA, van Velzen EJ, Gross G, Roger LC, Possemiers S, Smilde AK, Doré J, ve diğerleri. İnsan süperorganizmasındaki polifenollerin metabolik kaderi. Proc Natl Acad Sci. 2011;108(Ek 1):4531–8.MaddeGoogle Akademik
Price ND, Magis AT, Earls JC, Glusman G, Levy R, Lausted C, McDonald DT, Kusebauch U, Moss CL, Zhou Y, ve diğerleri. Kişisel, yoğun, dinamik veri bulutları kullanan 108 bireyden oluşan bir sağlık çalışması. Nat Biotechnol. 2017;35(8):747.MaddeCASGoogle Akademik
Lin Z, Zhang Q, Dai S, Gao X. Grup ve nükleer norm düzenlenmiş çok değişkenli regresyon yoluyla uzunlamasına hedefsiz metabolomik verilerde zamansal kalıpların keşfi. Metabolitler. 2020;10(1):33.MaddeCASGoogle Akademik
van de Pas NC, Woutersen RA, van Ommen B, Rietjens IM, de Graaf AA. İnsanlarda plazma kolesterol konsantrasyonlarını tahmin eden fizyolojik temelli bir siliko kinetik modeli. J Dudak Res. 2012;53(12):2734–46.MaddeGoogle Akademik
Adamko D, Rowe BH, Marrie T, Sykes BD, ve diğerleri. Normal insan idrarındaki metabolitlerin değişimi. Metabolomik. 2007;3(4):439–51.MaddeGoogle Akademik
Van Batenburg MF, Coulier L, van Eeuwijk F, Smilde AK, Westerhuis JA. Kapsamlı fonksiyonel genomik veriler için yeni değerler: metabolomik durum. Anal Kimya 2011;83(9):3267–74.MaddeGoogle Akademik
Smilde A, Westerhuis J, Hoefsloot H, Bijlsma S, Rubingh C, Vis D, Jellema R, Pijl H, Roelfsema F, Van Der Greef J. Dinamik metabolomik veri analizi: öğretici bir inceleme. Metabolomik. 2010;6(1):3–17.MaddeCASGoogle Akademik
Yamamoto H, Yamaji H, Abe Y, Harada K, Waluyo D, Fukusaki E, Kondo A, Ohno H, Fukuda H. Gizli değişkenlere farklı cezalar veren pca, pls, opls ve rfda kullanılarak metabolom verileri için boyutsallık indirgeme. Chemom Intell Lab Syst. 2009;98(2):136–42.MaddeCASGoogle Akademik
Nyamundanda G, Gormley IC, Brennan L. Uzunlamasına metabolomik verilerin analizi için dinamik olasılıklı ana bileşenler modeli. JR Stat Soc Ser C Appl Stat. 2014;63(5):763–82.MaddeGoogle Akademik
Smilde A, Bro R, Geladi P. Çok yönlü analiz: kimyasal bilimlerdeki uygulamalar. Chichester: Wiley; 2004.KitapGoogle Akademik
Acar E, Yener B. Gözetimsiz çok yönlü veri analizi: bir literatür taraması. IEEE Trans Knowl Data Eng. 2009;21(1):6–20.MaddeGoogle Akademik
Kolda TG, Bader BW. Tensör ayrıştırmaları ve uygulamaları. SIAM Rev. 2009;51(3):455–500.MaddeGoogle Akademik
Papalexakis EE, Faloutsos C, Sidiropoulos ND. Veri madenciliği ve veri füzyonu için tensörler: modeller, uygulamalar ve ölçeklenebilir algoritmalar. ACM Trans Intell Syst Technol. 2016;8(2):16.Google Akademik
Bader BW, Berry MW, Browne M. PARAFAC kullanılarak Enron e-postasında tartışma takibi. Londra: Springer; 2008. s. 147–63.
Dunlavy DM, Kolda TG, Acar E. Matris ve tensör faktörizasyonlarını kullanarak zamansal bağlantı tahmini. ACM TKDD. 2011;5(2):10.Google Akademik
Sun J, Papadimitriou S, Philip SY. Yüksek boyutlu ve çok yönlü akışlarda pencere tabanlı tensör analizi. Veri madenciliği üzerine altıncı uluslararası konferansta (ICDM’06). IEEE; 2006. s. 1076–80.
Acar E, Aykut-Bingol C, Bingol H, Bro R, Yener B. Epilepsi tensörlerinin çok yönlü analizi. Biyoenformatik. 2007;23(13):10–8.MaddeGoogle Akademik
Davidson I, Gilpin S, Carmichael O, Walker P. fMRI verilerinin kısıtlanmış tensör analizi yoluyla ağ keşfi. KDD’13: 19. ACM SIGKDD bilgi keşfi ve veri madenciliği uluslararası konferansının bildirileri. ACM; 2013. s. 194–202.
Roald M, Bhinge S, Jia C, Calhoun V, Adali T, Acar E. Parafac2 modelini kullanarak ağ evriminin izlenmesi. In: ICASSP’20: 45. IEEE akustik, konuşma ve sinyal işleme uluslararası konferansının bildirileri; 2020.
Yin K, Afshar A, Ho JC, Cheung WK, Zhang C, Sun J. Logpar: eksik değerlere sahip zamansal ikili veriler için lojistik parafac2 çarpanlarına ayırma. In: KDD’20: 26. ACM SIGKDD bilgi keşfi ve veri madenciliği uluslararası konferansının bildirileri; 2020.
Harshman RA. PARAFAC prosedürünün temelleri: “açıklayıcı” çok modlu faktör analizi için modeller ve koşullar. UCLA Work Pap Phonet. 1970;16:1–84.
Carroll JD, Chang JJ. “Eckart-young” ayrıştırmasının n-yollu genellemesi yoluyla çok boyutlu ölçeklemedeki bireysel farklılıkların analizi. Psychometrika. 1970;35:283–319.
Jansen JJ, Bro R, Hoefsloot HC, van den Berg FW, Westerhuis JA, Smilde AK. Parafasca: Asca, metabolik parmak izi verilerinin analizi için parafac ile birleştirildi. J Kemom. 2008;22(2):114–21.MaddeCASGoogle Akademik
van Heerden JH, Wortel MT, Bruggeman FJ, Heijnen JJ, Bollen YJ, Planqué R, Hulshof J, O’Toole TG, Wahl SA, Teusink B. Geçişte kayıp: glikolizin başlatılması, büyümeyen hücrelerin alt popülasyonlarını verir. Bilim. 2014;343:6174.
Tucker LR. Üç modlu faktör analizi üzerine bazı matematiksel notlar. Psychometrika. 1966;31(3):279–311.MaddeCASGoogle Akademik
Kruskal JB. Üç yollu diziler: üç doğrusal ayrıştırmaların sıralaması ve benzersizliği, aritmetik karmaşıklık ve istatistiklere uygulama. Doğrusal Cebir Uygulaması 1977;18(2):95–138.MaddeGoogle Akademik
Bro R, Harshman RA, Sidiropoulos ND, Lundy ME. Doğrusal olarak bağımlı yüklemelerle çok yönlü verilerin modellenmesi. J Chemom. 2009;23(7–8):324–40.MaddeCASGoogle Akademik
Schaefer U, Boos W, Takors R, Weuster-Botz D. Hücre içi metabolit dinamiklerini izlemek için otomatik örnekleme cihazı. Anal Biochem. 1999;270(1):88–96.MaddeCASGoogle Akademik
Hitchcock FL. Bir tensörün veya poliadiğin ürünlerin toplamı olarak ifadesi. J Math Phys. 1927;6(1–4):164–89.MaddeGoogle Akademik
Acar E, Dunlavy DM, Kolda TG, Mørup M. Eksik veriler için ölçeklenebilir tensör faktörizasyonları. Chemom Intell Lab Syst. 2011;106(1):41–56.MaddeCASGoogle Akademik
Tomasi G, Bro R. Parafac ve eksik değerler. Chemom Intell Lab Syst. 2005;75(2):163–80.MaddeCASGoogle Akademik
Kiers HA, Smilde AK. İkinci dereceden enstrümantal verilerle parametre tahmini için bir araç olarak kısıtlanmış üç modlu faktör analizi. J Chemom. 1998;12(2):125–47.MaddeCASGoogle Akademik
Bro R, Smilde AK. Bileşen analizinde merkezleme ve ölçekleme. J Chemom. 2003;17(1):16–33.MaddeCASGoogle Akademik
Bro R, Kiers HA. Parafak modellerindeki bileşen sayısını belirlemek için yeni ve etkili bir yöntem. J Chemom. 2003;17(5):274–86.MaddeCASGoogle Akademik
Stegeman A. Candecomp/parafac ve indscal’daki dejenerasyon, iki değerli tipik bir sıralamaya sahip birkaç üç dilimli dizi için açıklandı. Psychometrika. 2007;72(4):601–19.MaddeGoogle Akademik
Bro R. Parafac öğretici ve uygulamaları. Chemom Intell Lab Syst. 1997;38(2):149–72.MaddeCASGoogle Akademik
Acar E, Dunlavy DM, Kolda TG. Kanonik tensör ayrıştırmalarına uyum sağlamak için ölçeklenebilir bir optimizasyon yaklaşımı. J Chemom. 2011;25(2):67–86.MaddeCASGoogle Akademik
Bader BW, Kolda TG, ve diğerleri. Genel yazılım, son sürüm. MATLAB için Tensor Toolbox, Sürüm 3.1.
Andersson CA, Bro R. Matlab için n-yollu araç kutusu. Chemom Intell Lab Syst. 2000;52(1):1–4.MaddeCASGoogle Akademik
Wopereis S, Stroeve JH, Stafleu A, Bakker GC, Burggraaf J, van Erk MJ, Pellis L, Boessen R, Kardinaal AA, van Ommen B. Sağlıklı ve tip 2 diyabetik deneklerde standartlaştırılmış karışık yemek tolerans testinin çok parametreli karşılaştırması : phenflex mücadelesi. Genler Nutr. 2017;12(1):1–14.MaddeGoogle Akademik
Harshman RA. PARAFAC2: matematiksel ve teknik notlar. UCLA Çalışma Pap Phonet. 1972;22:30–47.Google Akademik
Bro R, Andersson CA. Çok yönlü algoritmaların hızının artırılması: Bölüm II: sıkıştırma. Chemom Intell Lab Syst. 1998;42(1–2):105–13.MaddeCASGoogle Akademik
Beutel A, Talukdar PP, Kumar A, Faloutsos C, Papalexakis EE, Xing EP. Flexifact: Hadoop’ta bağlı tensörlerin ölçeklenebilir esnek faktörizasyonu. In: 2014 SIAM uluslararası veri madenciliği konferansı bildirileri; 2014.
Jendoubi T, Ebbels TMD. Zaman seyri metabolik verilerinin ve biyobelirteç keşfinin bütünleştirici analizi. BMC Bioinform. 2020;21:11.MaddeGoogle Akademik
Acar E, Bro R, Smilde AK. Eşleştirilmiş matris ve tensör faktörizasyonlarını kullanarak metabolomikte veri birleştirme. IEEE Bildirileri. 2015;103:1602–20.MaddeCASGoogle Akademik
Teşekkürler
Amsterdam Üniversitesi Swammerdam Yaşam Bilimleri Enstitüsü’nden Dr. Meike T. Wortel’e, maya glikoliz modeli hakkındaki birçok yararlı iletişimi ve içgörüsü için teşekkür etmek istiyoruz. Ayrıca, Danimarka Pediatrik Astım Merkezi’ndeki (COPSAC) Dr. Morten Arendt Rasmussen ve işbirlikçilerimize de yararlı tartışmalar için minnettarız. Ayrıca, makalemizi iyileştirmemize yardımcı olan değerli yorumları için değerlendiricilere de minnettarız.
Finansman
Bu makalede sunulan çalışma Novo Nordisk Vakfı Hibe NNF19OC0057934 ve Norveç Araştırma Konseyi proje #300489 tarafından desteklenmiştir.
Stres kaynaklı dopamin D2 genindeki epigenetik metilasyonun ve nörolojik ve davranışsal sonuçlarının tanımlanması için yorumlar kapalı
D2 dopamin reseptörü (DRD2 ) geni, çeşitli nöropsikiyatrik bozukluklar arasında en kapsamlı çalışılan genlerden biri olarak önemli ilgi görmüştür. 1990 yılında özellikle DRD2 Taq A1 alelinin tanımlanması yoluyla şiddetli alkolizmle ilk ilişkisi kurulduğundan bu yana, farklı koşullardaki rolünü açıklamak için çok sayıda uluslararası araştırma yürütülmüştür. 22 Şubat 2024 itibarıyla PUBMED’de listelenen DRD2 genine odaklanan 5485 makale bulunmaktadır. Karışık sonuçlara sahip 120 meta-analiz yapılmıştır. Bizim görüşümüze göre, çeşitli DRD2 gen polimorfizmlerinin ilişkisiyle ilgili olumsuz raporların birincil nedeni, kontrollerin yetersiz taranması ve birçok gizli ödül eksikliği sendromu davranışının yeterince ortadan kaldırılmamasıdır. Ayrıca, DRD2 varyantlarının pleyotropik etkileri nöropsikolojik, nörofizyolojik, stres tepkisi, sosyal stres yenilgisi, anne yoksunluğu ve kumar bozukluğunda tanımlanmıştır ve epigenetik DNA metilasyonu ve histon translasyon sonrası negatif metilasyonu bu makalede tartışıldığı gibi tanımlanmıştır. 19 Ekim 2022 itibarıyla PUBMED’de DNA metilasyonu için 70 makale ve histon metilasyonu için 20 makale listelenmiştir. Bu yorum için DNA ve/veya histon metilasyonunu belirtmedik; bunun yerine davranışsal etkilere dayalı kısa bir özet sağladık. Blum ve Noble’ın DRD2 Taq A1 alelini genelleştirilmiş bir ödül geni olarak nitelendirdiği ve mutlaka spesifik alkolizm olmadığı gerçeğine dayanarak, artık alanın nöroepigenetik hakaretleri düzenlemek için efektör grupları kullanmanın veya muhtemelen “dopamin homeostazisini” indükleyerek negatif mRNA azaltılmış ifadeyi potansiyel olarak ortadan kaldırma fikrini kullanmanın yollarını bulması gerekmektedir.
Blum’un araştırma ekibi 2013 yılında alkol tüketiminin ve özellikle stresli durumlara yanıt olarak yüksek oranda uyuşturucu arama davranışlarının ardındaki motivasyonları araştırmaya başladı. 1 – 4 Amaçları, bu tür davranışları yönlendiren altta yatan mekanizmalar hakkında farkındalık yaratmak ve anlayış geliştirmekti. Dahası, yüksek riskli yenilik durumlarını arayan ve bunlara katılan milyonlarca bireyle ilgili ek sorular sordular ve haz arayan davranışların sonuçlarını düşündüler. Bu soruların cevaplarının beyinlerimizin karmaşık işleyişinde ve belki de genetik yapımızda yatabileceğini öne sürdüler. 5 , 6
Amerika şu anda tarihin en kötü opioid salgınıyla karşı karşıya. Hastalık kontrol ve önleme merkezleri (CDC), 2021’de ABD’de 106.699 aşırı doz ölümünü bildirdi ve bu ölümlerin 80.441’i opioidlerle ilgiliydi. Ayrıca, CDC’den gelen geçici veriler, 2022’de opioid ölümlerinin yaklaşık 82.998’e yükseleceğini gösterdi. 7 , 8
Blum ve arkadaşları , 1995 yılında bağımlılık ve dürtüsel davranışların potansiyel bir öngörücüsü olarak ödül eksikliği sendromu (RDS) kavramını ortaya koydular. 5 , 9 – 11 Bu teori o zamandan beri çok sayıda çalışma tarafından desteklendi ve RDS de dahil olmak üzere psikiyatrik bozukluklarda gözlemlenen davranışsal, bilişsel ve duygusal bozuklukların nörolojik yollardaki işlevsel eksikliklerle bağlantılı olduğunu gösterdi. 12 – 17 Örneğin, D2 dopamin reseptörü ( DRD2 ) Taq A1 aleli, saldırganlık, 18 alkolizm, bağımlılık davranışları 19 ve nöropsikiyatrik bozukluklar dahil olmak üzere çok sayıda davranışsal fenotiple tutarlı bir şekilde ilişkilendirilmiştir . Hem DRD1 hem de DRD2 genleri ödül yolları ve mekanizmalarıyla bağlantılıdır. 20 Mezolimbik bölgelerdeki biyokimyasal süreçlerin doruk noktasına ulaşması, özellikle sinaptik boşluklardaki artan dopamin seviyelerinin DRD1 , DRD2 ve diğer reseptör alt tipleriyle etkileşime girdiği nucleus accumbens’te (NAc) ödüllendirici fenomenlere yol açar . 21 , 22 Pozitron emisyon tomografisi çalışmaları , esrar, psikostimülan, opioid veya alkole bağımlı olan bireylerde ve obez olanlarda kontrol deneklerine kıyasla daha düşük DRD2 bulunabilirliğini göstererek daha fazla içgörü sağlamıştır . 23 – 28
Ayrıca, çalışmalar DRD2 geninin A1 ve B1 minör alelleri ile kokain kullanım bozukluğu (CUD) arasında bir ilişki olduğunu ortaya koymuştur. 26 Bu bulgular, kromozom 11’deki q22-q23 bölgesinde bulunan DRD2 genindeki genetik varyasyonların psikostimülan kullanım bozukluğuna (PUD) karşı artan duyarlılığa katkıda bulunduğunu göstermektedir. İlginç bir şekilde, bu gözlemler, kokain kötüye kullanımı ve bağımlılığıyla mücadelede D2 agonisti olan bromokriptinin potansiyel terapötik etkinliğine işaret eden Gold’un grubunun erken araştırmalarıyla örtüşmektedir. 29 D2 reseptör seviyelerinin azalmasıyla ilişkili olan DRD2 Taq A1 alelinin aksine , ana varyant olan DRD2 Taq A2 aleli, normal D2 reseptör seviyeleriyle karakterizedir ve muhtemelen psikostimülan kötüye kullanımı ve istismarına karşı koruma sağlar. 30
Tarihsel olarak, Dackis ve Gold, güçlü D2 agonisti bromokriptinin şiddetli kokain bağımlılığı için epigenetik tedavi olarak kullanımını öneren ilk kişiler arasındaydı. 29 Ancak klinik çalışmalar, bromokriptinin DRD2 reseptörlerinin aşağı düzenlenmesine yol açtığını ve bu amaç için etkili olmadığını ortaya koydu; bu da sınırlı klinik kullanımına yol açtı. 31
Bu yorum , hem maddeyle ilişkili hem de maddeyle ilişkili olmayan RDS davranışlarında DRD2 geninde meydana gelen epigenetik değişiklikleri tanımlayan çok sayıda çalışmayı ortaya çıkarmıştır .
2. Epigenetik ve bağımlılık davranışları
Epigenetik, hücre çekirdeğindeki kromatine yüklenen moleküler modifikasyonları ifade eder ve kromatin organizasyonu, DNA onarımı, RNA transkripsiyonu ve ekleme gibi diğer temel hücresel işlevler dahil olmak üzere çeşitli DNA ile ilgili süreçleri düzenlemede önemli bir rol oynar. 32 Madde kullanım bozukluğu (SUD), çevresel faktörlerin gen ifadesini nasıl etkileyebileceğinin başlıca bir örneğidir. 33 Bu bağlamda, bir kişinin deneyimleri, özellikle istemli tekrarlayan uyuşturucu kullanımı, beyindeki epigenomu belirli beyin bölgelerine ve hücre tiplerine özgü bir şekilde değiştirebilir. 34 İlaçların neden olduğu DNA ile ilgili süreçlerin düzensizliği ve modifikasyonunun, psikoaktif madde bağımlılığının patogenezini kolaylaştıran anormal hücresel işlevlere katkıda bulunabilecek epigenetik değişikliklere yol açabileceği varsayılmaktadır. Nestler’in grubu daha önce epigenetik süreçlere ilişkin içgörüler edinmenin terapötik açıdan umut vaat ettiğini öne sürmüştü. Beyindeki önemli uyuşturucu kaynaklı epigenetik değişiklikleri hedeflemek, uyuşturucu bağımlılığı döngüsünü bozabilir ve böylece bireylerin amansız bir bağımlılık döngüsüne yenik düşmesini önleyebilir .
Nöroepigenetik manzaranın karmaşıklıklarını anlamak, epigenetik modifikasyonların dizisini kabul etmeyi gerektirir. 35 Bunlar arasında, olumsuz ortamlarda meydana gelen önemli bir epigenetik değişim, metilasyon ve hatta dopaminilasyon gibi histon translasyon sonrası modifikasyonlarını (PTM’ler) içerir. 36 , 37 Esasen, kromatin, histon protein oktamerlerinin etrafına karmaşık bir şekilde sarılmış ve nükleozomlar oluşturan DNA’dan oluşur ve bu da daha sonra hücre çekirdeği içinde kompakt bir paketlemeye olanak tanır. Bu yapısal düzenleme, dış uyaranlara yanıt veren uyarlanabilir bir iskele görevi görür. Özellikle, histonlar arginin ve lizin kalıntıları açısından zengindir ve bu da oldukça temel yapılarına katkıda bulunur. Nükleozom çekirdeğinden uzanan histon N-terminal kuyruklarındaki bu kalıntıların ve diğerlerinin PTM’leri, kromatinin fiziksel özelliklerini ve yük dağılımını düzenleyerek DNA ile ilişkili süreçleri düzenler.
Histon alt birimleri, asetilasyon, metilasyon, fosforilasyon, adenozin difosfat ribozilasyonu, ubikitinasyon ve sumoyilasyon dahil ancak bunlarla sınırlı olmamak üzere çok sayıda PTM’ye maruz kalır ve yeni tanımlanan modifikasyonların genişleyen dizisi vardır. 38 , 39 Bu modifikasyonlar histon proteinleri boyunca 50’den fazla farklı bölgede meydana gelir. 38 , 39
Allis ve diğerlerine göre , “histon PTM’leri geri dönüşümlüdür; “yazar” enzimler tarafından dinamik olarak biriktirilir, hücresel yanıtı aracılık eden “okuyucu” proteinler tarafından tanınır ve “silgi” enzimleri tarafından uzaklaştırılır.” 36 Çalışmalar, çok sayıda yazar, silgi ve okuyucu proteinin ifadesinin ve aktivitesinin hem bağımlı bireylerde hem de bağımlılığın hayvan modellerinde düzensiz olduğunu göstermiştir. 40 – 42 Bu düzensizlik, bağımlılık için yeni epigenetik tedavilerin geliştirilmesine olan ilgiyi artırmıştır. Bu proteinlerin normal işlevlerinin, dopamin gibi nörotransmitter seviyelerini yeniden dengelemeye yardımcı olan küçük molekül müdahaleleri veya işlevsel gıda kompleksleri yoluyla yeniden kazandırılması, bağımlılık karşıtı epigenetik tedaviler için umut verici bir yol sunmaktadır. 43 – 52
3. Epigenetik biyobelirteçler
3.1. DNA metilasyonu
Biyobelirteç, biyolojik bir molekülün, durumun veya koşulun niceliksel, ölçülebilir bir göstergesidir. 53 DNA metilasyonu, kimyasal kararlılığı, memeli gelişimi ve hastalığındaki rolü ve çok çeşitli biyolojik süreçlerde gen ifadesini düzenlemedeki önemli rolü nedeniyle en çok araştırılan epigenetik biyobelirteçtir. 54 , 55 DNA metilasyonu gen yapısında fiziksel değişikliklere neden olsa da doğası gereği geri dönüşümlüdür. Birincil işlevi, DNA transkripsiyonunu engellemek ve böylece belirli genlerin ifadesini bastırmaktır. DNA metilasyon testleri daha uygun fiyatlı ve erişilebilir hale gelmiştir ve vücut sıvılarından, hücrelerden veya dokulardan elde edilebilen yalnızca az miktarda DNA gerektirir. 19 Ayrıca, DNA bakteri, virüs, bitki veya memelilerden izole edilebilir. DNA metilasyonu, DNA moleküllerine bir metil grubunun eklenmesiyle karakterize edilen bir biyokimyasal işlemdir. Yaygın bir durum, bir sitozin halkasının 5 karbon pozisyonuna bir metil grubunun eklenmesi ve 5-metilsitozin (5-mC) oluşturulmasıdır. 56 DNA metilasyon analizleri, DNA örnekleri içindeki 5-mC seviyelerini ölçmek için kullanılan tekniklerdir. DNA metiltransferaz (DNMT) enzimi, özellikle CpG dinükleotid bölgelerinde DNA metilasyonunu katalize etmede önemli bir rol oynar. 57 Özellikle, DNMT-1, vücutta en yüksek DNA metilasyonu seviyelerini gösteren beynin mitotik hücrelerinde DNA replikasyonundan birincil olarak sorumludur. 58 Öte yandan DNMT-3A ve DNMT-3B, erken gelişim aşamalarında metilasyon modellerini düzenler. 59
NA metilasyonu erken beyin gelişiminde ve gen ifadesi yoluyla bölgelerin belirlenmesinde kritik bir rol oynar. 57 Ayrıca çeşitli kanserlerle ilişkili mutasyon olaylarını önemli ölçüde etkileyerek gen mutasyonları ve spesifik tümör baskılayıcı genlerin inaktivasyon riskini artırır. 60 – 64 Örneğin, kirlilik gibi çevresel kanserojenlere maruz kalma, DNA metilasyonundan sorumlu genlerde mutasyonlara neden olabilir ve bu da çoğalma veya farklılaşma gibi değişmiş hücresel durumlara ve sonuçta kansere yol açabilir. 64
Üstelik, DNA metilasyonu, genlerin DNA metilasyonu yoluyla susturulduğu veya etkisizleştirildiği bir süreç olan genomik baskılamada yer alır. Baskılama, babadan veya anneden gelen bir alel susturulduğunda ve bunun sonucunda baskılanmış bir gen oluştuğunda meydana gelir. 65 Bu köken ebeveyn etkileri gametler tarafından kalıtılabilir ve yavrulara aktarılabilir ve Prader-Willi sendromu ve Angelman sendromu gibi çeşitli hastalıklara yol açabilir. 66 , 67 Bu kalıtım şekli ayrıca majör depresif bozukluk, bipolar bozukluk, şizofreni, otizm ve ilgili durumlar gibi psikiyatrik bozukluklarda DNA metilasyonunun rolünün anlaşılmasına yardımcı olur. Bu bulgu önemlidir çünkü bu mekanizmadaki anormalliklerin çeşitli bozuklukların patofizyolojisine nasıl katkıda bulunduğuna dair önemli içgörüler sağlar ve DNA metilasyonunun terapötik bir hedef olarak potansiyelini gösterir. 67
3.2. DNA demetilasyon
DNA demetilasyon, DNA metilasyonu ile birlikte gerçekleşen ancak yaygın olarak anlaşılmayan bir işlemdir. 68 DNA hasarı için bir biyobelirteç görevi görür ve hem bölünen hem de bölünmeyen hücrelerde aktif veya pasif olarak meydana gelebilen DNA’dan bir metil grubunun çıkarılmasını içerir. Pasif demetilasyon, DNA replikasyonu sırasında 5mC’nin kaybını gerektirirken, aktif demetilasyon 5 mC’den bir metil grubunun değiştirilmesini veya çıkarılmasını içerir. Özellikle, sitozinin metillenmiş bir formu olan 5 mC, gen mutasyonları için bir ilgi noktası ve gen transkripsiyonundaki düzenleyici rolü nedeniyle bir epigenetik belirteç olarak yaygın olarak kullanılır. 55 , 69
Ayrıca, 5-hidroksimetilsitozin (5hmC) olarak bilinen 5mC’nin bir türevi, özellikle beyin olmak üzere çeşitli organ dokularında bol miktarda bulunur. DNA demetilasyon, DNA hasarı için bir belirteç görevi görür ve potansiyel olarak mutasyona uğramış bölgeleri belirleyerek onarım süreçlerini kolaylaştırır. 70 Bir embriyonun ve gelişmekte olan zigotların primordial germ hücreleri, demetilasyonun meydana geldiği birincil bölgelerdir ve bu da farklılaşma mekanizmalarındaki temel rolünü vurgular. 71
DNA demetilasyonuna on-on bir translokasyon (TET) ailesinden enzimler aracılık eder. 72 , 73 TET enzimleri çeşitli malignitelerde tümör baskılayıcı olarak işlev görür ve bunların kaybı veya işlev bozukluğu hızla mutasyona uğrayan kanserlerle yakından ilişkilidir. 74 Ek olarak, timin DNA glikozilazı, hücre döngüsü boyunca hasarlı DNA’nın onarımı için gerekli olan baz eksizyon onarımını başlatarak DNA demetilasyonunda ve normal gelişimde önemli bir rol oynar. 75
Özetle, DNA demetilasyonunun çevresel risk faktörlerinden (örneğin yaralanmalar ve madde kullanımı) etkilenen genlerin epigenetik yeniden programlanmasında bir mekanizma olarak görev yaptığı görülmektedir.
3.3. Histon modifikasyonu
Bir protein sınıfı olan histonlar, DNA’nın nükleozom olarak bilinen yapısal birimlere paketlenmesini kolaylaştırır. Bu paketleme yapısal destek sağlar ve DNA’nın hücre çekirdeğine uygun şekilde yerleşmesini sağlar. Histonların modifikasyonu, transkripsiyonu etkilediği ve çevresel uyaranlara ve stres faktörlerine yanıt olarak fenotipleri değiştirdiği için epigenetik düzenlemeye örnek teşkil eder. Histon modifikasyonları, DNA’yı sıkıca sıkıştırarak ve hücresel makineye erişimini engelleyerek bu süreci başarır. 76 Tersine, histon gevşemesi, proteinlerin DNA’ya erişimini artırarak hücre tarafından analiz edilmeye olan duyarlılığını artırır. Histon değişikliklerinin, hücre durumu değişikliklerine ek olarak DNA onarımını ve replikasyonunu da etkilediği bulunmuştur. 77 Ayrıca, lipitler ve karbonhidratlar gibi makromolekülleri sentezlemek ve hücre metabolizmasını ve enerji çıktılarını düzenlemek için kullanılırlar. 78 Histon değişikliklerinin sıklıkla proteinler aracılığıyla aşağı akış sinyalleme kaskadlarını aktive ederek dolaylı bir etkiye sahip olduğunu belirtmek önemlidir.
3.4. Histon asetilasyonu
Histon asetilasyonu, genellikle gelişmiş gen transkripsiyonu ile ilişkilendirilen ve hafıza oluşumu ve uyuşturucu bağımlılığının altında yatan süreçlerde rol oynayan geri dönüşümlü bir epigenetik mekanizmayı temsil eder. Özellikle uyuşturucu bağımlılığından kaynaklanan çevresel ipuçlarının gen ifadesinde hedeflenen değişiklikleri ortaya çıkarmak için bir kanal görevi görür. Asetilasyonun enzimatik düzenlenmesi, histon deasetilazları (HDAC’ler) ve histon asetiltransferazları (HAT’ler) içerir. 79 Histon asetilasyonu sırasında, negatif yüklü bir asetil grubu histon proteinlerindeki lizin kalıntılarına bağlanır ve 80 transkripsiyon faktörü bağlanmasına ve artan gen ifadesine elverişli gevşemiş bir kromatin yapısıyla sonuçlanır.
Histonlar arasında H3 ve H4 oldukça korunmuştur ve kromatin organizasyonundaki temel rolleri nedeniyle önemli ilgi görmüştür. 81 Histon asetilasyonu genellikle diğer kromatin modifikasyonlarıyla birlikte meydana gelir. DNA metilasyonu ve demetilasyonuna benzer şekilde, histon asetilasyonunun çeşitli hastalıklar için bir biyobelirteç olarak işlev görmesi beklenmektedir. Histon asetilasyonundaki azalma nörogelişim bozuklukları, nöral dejenerasyon, plastisite ve hafıza bozukluğu ile ilişkilendirilmiştir. CLOCK proteini HAT’ı örneklendirir ve sirkadiyen ritimler ve hücresel homeostazın ayrılmaz bir parçası olan önemli bir transkripsiyon faktörü olarak durmaktadır. Bu HAT’taki bozulmalar uyku yoksunluğu ve bipolar bozukluk manik benzeri davranışlarla ilişkili bir dizi değişikliğe neden olur. 82 Histonların kapsamlı bir şekilde araştırılması, psikiyatrik bozukluklar, nörodejeneratif hastalıklar ve tümör hücreleri için çok sayıda terapötik stratejinin geliştirilmesi için önemli olacaktır.
3.5. Kromatin yeniden şekillenmesi
Histon proteinlerindeki değişiklikler, asetilasyon, deasetilasyon, metilasyon ve demetilasyon gibi değişiklikleri kapsayan kromatinin yapısını etkileyebilir. 83 Bu değişiklikler nihayetinde transkripsiyonun aktive mi yoksa baskılanmış mı olacağını belirler. Sıkıca yoğunlaşmış ve transkripsiyonel olarak inaktif olan kromatine heterokromatin denir ve burada genler tipik olarak susturulur veya inaktif hale getirilir. Tersine, ökromatin, transkripsiyon için daha erişilebilir olan gevşekçe yoğunlaşmış kromatini ifade eder. Ökromatinde, DNA transkripsiyon faktörleri ve diğer DNA bağlayıcı proteinler tarafından bağlanmak için kolayca erişilebilirdir ve bu da gen ifadesinin düzenlenmesini kolaylaştırır. 84 Kromatinin yoğunlaşmış bir durumdan daha erişilebilir bir duruma geçiş süreci, histon değişikliklerini ayrıntılı olarak açıklayan önceki bölümlerde tartışıldığı gibi, kromatin yeniden şekillenmesi olarak adlandırılır.
Kromatin, DNA onarımı, DNA replikasyonu, kromozom ayrımı ve sinyal iletimi gibi çok sayıda hücresel işlevde önemli bir rol oynar. Kromatin yeniden düzenleyicileri tipik olarak histonları modifiye etmek ve nükleozomları yeniden yapılandırmak için adenozin trifosfat hidrolizini kullanan çok proteinli kompleksler ve enzimlerden oluşur. Kromatin yeniden düzenlemesinin bir örneği embriyonik bir kalbin gelişiminde belirgindir. Bu yeniden düzenleyicilerden kaynaklanan mutasyonlar veya kusurlar yetişkinlerde çeşitli kardiyovasküler bozukluklara yol açabilir. 85
3.6. Kodlamayan RNA (ncRNA) ekspresyonu
ncRNA, DNA’dan transkripsiyona uğrayan ancak proteinlere çevrilmeyen RNA moleküllerini ifade eder. Gen ekspresyonunu düzenlemede rol oynayan dört ana ncRNA türü vardır: MikroRNA’lar (miRNA), uzun ncRNA’lar (lncRNA), Piwi ile etkileşen RNA’lar ve kısa karışan RNA’lar. Bunların hepsi kapsamlı bir şekilde araştırılmıştır. 86 Bu düzenleyici mekanizmalar, transkripsiyon ve translasyonu yönetmede, kromatin yeniden şekillenmesini etkilemede önemli roller oynar ve fizyolojik süreçlerin yanı sıra hastalık durumlarının da ayrılmaz bir parçasıdır. 87 MiRNA’lar, hedef RNA’lara bağlanarak gen ekspresyonunu baskılayarak ve translasyonu inhibe ederek işlev görür. Genetik bilginin hücreler içinde, farklı hücreler ve dokular arasında ve hatta anne sütü, ter, idrar ve kan gibi vücut sıvıları arasında aktarılmasını kolaylaştırırlar. Kardiyovasküler bozukluklarda ve tümör baskılanmasında rolleri nedeniyle miRNA’lar mükemmel tanı belirteçleri olarak kabul edilirler. 60 , 64 Buna karşılık, lncRNA’lar daha hücre tipi spesifiktir ve miRNA’lara kıyasla daha düşük seviyelerde ifade edilirler. Bunlar öncelikle çekirdeğin içinde veya dışında meydana gelebilecek transkripsiyonu düzenler. Ek olarak, lncRNA’lar bir hücrenin sitoplazması içindeki mRNA’nın düzenlenmesi de dahil olmak üzere çeşitli işlevler sergiler. 86
4. RDS için terapötik hedef olarak DRD2 geni
Kör bir çalışmada Blum ve arkadaşları , 88 DRD2 geninin alkolizmle ilk alelik ilişkisini gösterdiler . Çalışmada hem alkolik olmayanlardan hem de alkolizm teşhisi konmuş bireylerden toplanan 70 beyin örneği kullanıldı. DNA örnekleri restriksiyon endonükleazlarıyla sindirime tabi tutuldu ve tam 3′ kodlama ekzonu, poliadenilasyon sinyali ve insan DRD2 geninin ( lambda hD2G1) yaklaşık 16,4 kilobaz (kb) kodlamayan 3′ dizisini içeren bir klonla araştırıldı. Bulgular, DRD2 geninin A1 alelinin varlığının alkolizmli bireylerin %77’sini doğru bir şekilde sınıflandırdığını, yokluğunun ise alkolik olmayanların %72’sini doğru bir şekilde sınıflandırdığını ortaya koydu. DRD2 geninin bu polimorfik deseni, kromozom 11’in q22-q23 bölgesinde en az bir alkolizm türü için bir duyarlılık geninin varlığını önerdi.
Benzer beyin dokusunu kullanarak, Noble ve arkadaşları 23 hem alkolik hem de alkolsüz deneklerden alınan 66 beyinde , insan DRD2 geninin DRD2 reseptörünün bağlanma özellikleriyle alel ilişkisini araştıran bir çalışma yürüttüler. Bu kör deneyde, serebral korteksten çıkarılan DNA, restriksiyon endonükleaz Taq ile işleme tabi tutuldu ve insan DRD2 geninin bir klonunun (XhD2G1) 1,5 kb’lik bir özetiyle araştırıldı . Bağlanma afinitesi (K d ) ve bağlanma yeri sayısı (B max ) dahil olmak üzere DRD2’nin bağlanma özellikleri, ligand olarak tritiyumlanmış spireron kullanılarak bu beyinlerin kaudat çekirdeklerinde belirlendi. Sonuçlar, alkolik olmayan deneklerle karşılaştırıldığında, ayarlanmış K d’nin alkolik deneklerde önemli ölçüde daha düşük olduğunu, bunun da ikinci grupta DRD2’nin daha yüksek bağlanma afinitesine sahip olduğunu gösterdiğini ortaya koydu. Ek olarak, alkolizmle güçlü bir ilişki gösteren A1 alelini taşıyan deneklerde B max’ın daha düşük olduğu bulunurken, A2 alelini taşıyan deneklerde B max’ta önemli bir değişiklik gözlenmedi. Dahası, A2/A2, A1/A2 ve A1/A1 alellerine sahip deneklerde giderek azalan bir B max bulundu; A2/A2 alellerini taşıyan deneklerde en yüksek ortalama değerler, A1/A1 alellerini taşıyanlarda ise en düşük ortalama değerler görüldü. DRD2 geninin gözlemlenen polimorfik örüntüsü ve reseptör ifadesindeki varyasyonu, dopaminerjik sistemin bireyleri en az bir şiddetli alkolizm alt tipine yatkın hale getirmede rol oynadığını güçlü bir şekilde düşündürmektedir. Daha sonraki çalışmalar bu bulguları doğrulamıştır, 89 – 104 ve 22 Şubat 2024 itibarıyla, “Dopamin D2 Reseptör Geni” terimini kullanan bir PubMed araması toplam 5485 listeleme sağlamıştır.
Kanabinoiderjik, kolinerjik, serotonerjik, opioiderjik, glukonergik, GABAerjik ve glutaminerjik olmak üzere en az yedi ana nörotransmitter yolunu etkileyen epigenetik hakaretlerin farkında olsak da, odak noktamız dopaminerjik sistem, özellikle DRD2 geni üzerindedir. “Metilleme ve Dopamin D2 Reseptör Geni” arama terimini kullanan PUBMED listelemelerine dayanan birincil inceleme, 22 Şubat 2024 itibarıyla 378 listeleme üretti.
5. Epigenetik modifikasyonlar ve DRD2 geni
Birçok ödül geninin ve ilişkili lokusların uyuşturucu madde (yani alkol) tüketiminden sonra epigenetik değişikliklere uğradığının gösterildiğini kabul ederken, bu yorumun amacı epigenetik değişiklikleri, özellikle DRD2 geninde tanımlamaya odaklanan bir anlatı sunmaktır . Bu yorumda, DNA ve/veya histon metilasyonunu belirtmeyi tercih etmedik, bunun yerine davranışsal etkilere dayalı bir özet sunuyoruz.
Bohnsack ve ark . tarafından yakın zamanda yapılan bir çalışma, istenmeyen davranışları zayıflatmak için epigenetik histon modifikasyonlarını hedefleme ve düzenlemeye dair bir örnek sunmaktadır. 104 Ergenlik döneminde aşırı alkol tüketiminin, aktiviteyle düzenlenen sitoskeletonla ilişkili protein (ARC) acil-erken geninin güçlendirici bölgesinde epigenetik modifikasyonlara neden olduğu yaygın olarak bilinmektedir; bu gen özellikle sinaptik aktivite yanıt elementi (SARE) olarak bilinir. 104 Bu epigenetik modifikasyonlar , hem kemirgen modellerinde hem de insan çalışmalarında gözlemlenen bir fenomen olan amigdaladaki ARC ekspresyonunda bir azalmaya yol açar . 104
Pandey’in grubu tarafından yürütülen bir deneyde, dCas9-P300 kullanımının ARC SARE’de histon asetilasyonunu artırdığı gösterildi . Bu müdahale, ARC ifadesindeki eksiklikleri etkili bir şekilde normalleştirdi ve sonuç olarak ergenlik döneminde alkol maruziyetini inceleyen bir sıçan modelinde yetişkin anksiyetesinde ve aşırı alkol tüketiminde azalmaya yol açtı. 104 İlginç bir şekilde, dCas9-Kruppel ile ilişkili kutunun (KRAB) aksine, ARC SARE’de baskılayıcı histon metilasyonunu teşvik ettiği, bunun sonucunda ARC ifadesinde azalma, anksiyete benzeri davranışların ortaya çıkması ve kontrol grubunda alkol tüketiminde artış olduğu bulundu .
5.1. SÜD
Epigenetik SUD’a odaklanan çalışmaların bir özeti sunulmaktadırtablo 1SUD’un ayrıntılı açıklamaları aşağıdaki alt bölümlerde verilmiştir.
Tablo 1.
Madde kullanım bozukluğunun epigenetiği
Madde türü
Bulgular
Referanslar
Kenevir
Aile geçmişi olan DRD2 geninde daha yüksek histon metilasyonu
METH ile tedavi edilen farelerde ayrıca (i) toplam H3ac ve H4ac’nin genel düzeylerinde azalma ve 5-metilsitozin (5-mC)’nin genel düzeylerinde artış ve (ii) DRD2 promotörlerinde H3ac zenginleşmesinde azalma görüldü
Hem insan hem de hayvan kendi kendine uygulama deneylerinde, histon işaret proteini-R-metiltransferaz-6 (PRMT6) ve histon H3 üzerindeki R2’nin asimetrik demetilasyonunun (H3R2me2a) kemirgen ve kokain bağımlı insan NAc’de azaldığı görüldü.
METH, METH psikozu olan hastalarda dopaminle ilişkili bir dizi genin (yani DRD3, DRD4 ve COMT) DNA metilasyonunda azalmaya neden olurken, METH psikozu olmayan hastalarda bu azalmaya neden olmaz.
Oyaci ve arkadaşları tarafından yürütülen bir çalışmada , 105 DRD2 genindeki DNA metilasyon seviyeleri, hastalar arasında klinik parametrelere ve DRD2 genotip dağılımına göre karşılaştırıldı . Bulguları, özellikle CUD veya sentetik kanabinoid kullanım bozukluğu (SCUD) aile öyküsünün varlığı ile ilgili olarak, gruplar arasında metilasyon durumunda önemli farklılıklar ortaya koydu. Çalışmanın dikkate değer bir sınırlaması, yanlış kontrollerin kullanılmasıdır (yazarlar kontrolleri kumar oynama ve aşırı yeme gibi diğer potansiyel bağımlılık davranışları açısından taramamıştır). Böyle bir tarama yapılmadan, örneğin DRD2 A1 alelinin varlığının %40 kadar yüksek olabileceği bilinmektedir. Bu kusur, araştırmacıların hem CUD hem de SCUD ile yüksek DNA metilasyonu için doğrudan kanıt oluşturmasını engellemiş olabilir. Gerra ve arkadaşları tarafından yürütülen ayrı bir çalışmada , 106 DRD2-ANKKI gen bölgesindeki DNA metilasyonunu araştırmak için hem CUD hastalarını hem de kontrol gruplarını içeren genetik ve sosyodemografik bir analiz yapıldı . DRD2 geninin 8. eksonunda önemli hipermetilasyon tespit ettiler . İlginç bir şekilde, çalışma ayrıca daha yüksek eğitim seviyeleri ile CUD riskinin azalması arasında bir korelasyon ortaya çıkardı. Yazarlar, DRD2 geninin 8. eksonunda farklı şekilde metillenmiş bölgeler (DMR’ler) bulgularının farmakolojik terapötik ajanların geliştirilmesi için potansiyel biyobelirteçler veya hedefler olarak hizmet edebileceğini akıllıca öne sürdüler.
DiNieri ve ark . tarafından 107 , delta-9-tetrahidrokanabinol (THC)’e doğum öncesi maruziyetin bir hayvan modelini kullanan bir çalışmada , doğum öncesi kanabis maruziyetinin , beyindeki önemli bir ödül bölgesi olan NAc’de DRD2 haberci RNA ifadesinde bir azalmaya yol açtığı gözlemlendi. Dahası, Hurd’un grubu 107 , THC’ye maruz kalan gebe sıçanların yavrularında histon metilasyon farklılıklarını tanımladı. Yetişkin NAc’nin kromatin immünopresipitasyonu, THC’ye maruz kalan yavrularda 2meH3K9 baskılayıcı işaretinde bir artış ve DRD2 gen lokusunda 3meH3K4 ve RNA polimeraz II’de bir azalma ortaya koydu. Bu azalmış DRD2 ifadesi, azalmış DRD2 bağlanma bölgeleri ve yetişkinlikte opiat ödülüne karşı artan duyarlılıkla ilişkilidir . Dahası, çalışma maternal kanabis kullanımının histon lizin metilasyonunu yöneten epigenetik mekanizmaları değiştirdiğini ve böylece yavrularda mezolimbik DRD2’nin gelişimsel düzenlenmesini etkilediğini ileri sürmüştür. DRD2’deki bu sonraki azalma , yavruların gelecekte bağımlılığa yatkınlığını artırabilir. İlgi çekici olarak, Blum’un laboratuvarından 108 daha önceki bir çalışma, DiNieri ve diğerlerinin bulgularıyla uyumlu olarak, THC’ye doğum öncesi maruziyetten kaynaklanan vas deferenslerde önemli bir değişiklik olduğunu ve enkefalinlere karşı artan bir duyarlılık olduğunu göstermiştir . 107
5.1.2. Tütün
Liu ve ark . tarafından vurgulandığı gibi , 109 önceki araştırma, Avrupa ve Afrika kökenli bireylerde bağımlılık duyarlılığında NCAM1-TTC12-ANKK1-DRD2 gen kümesinin önemini vurgulamıştır . Bu temel üzerine inşa edilen Liu ve meslektaşları, sigarayla ilişkili DMR’leri ortaya çıkarmak için yeni nesil bisülfit dizilimi gerçekleştirdiler. Haplotip tabanlı ilişki analizi yoluyla, günlük sigara sayısı ile rs4245148, rs4581480, rs4648317 ve rs11214613 tarafından oluşturulan DRD2’deki CTAG haplotipi arasında önemli bir ilişki belirlediler . Dahası, çalışma, üçü DRD2/ANKK1 bölgesinde bulunan dört önemli sigarayla ilişkili DMR belirledi. Bu bulgular, Çinli erkek sigara içicileri arasında nikotin bağımlılığı ile ANKK1/DRD2 bölgesindeki varyantlar ve haplotipler arasında dikkate değer bir ilişki olduğunu açıklamaktadır . Daha da önemlisi, sonuçlar bu tür ilişkileri kolaylaştırmada DNA metilasyonunun oynadığı önemli rolün de altını çiziyor.
5.1.3. Alkol
DRD2 bağlanma potansiyeli ile ödüllendirici uyaranlara ve madde kullanımına verilen nöral tepkiler arasında güçlü korelasyonlar gözlemlenmiştir. 89 – 104 Sonuç olarak, DRD2 işlevlerindeki bozulmalar, madde bağımlılığının patofizyolojisini açıklayan teorik modellerin kritik bir bileşenini oluşturur. 110 Dahası, Pandey ve ark . tarafından önerildiği gibi, 111 epigenetik modifikasyonlar, alkol maruziyetinin beyni nasıl etkilediğini etkileyen temel moleküler mekanizmalar olarak hizmet edebilir.
Bu fikir , yaş kontrol edildikten sonra DRD2 promotör DNA metilasyonunun sol kaudat, sağ kaudat, sol putamen, sağ putamen ve sağ Nac’deki alkol ipuçlarına verilen yanıtlarla pozitif ilişkili olduğunu gösteren Bidwell ve arkadaşları tarafından araştırılmıştır . 77 , 112 Bu bulgu, ödül ipuçlarına yanıt olarak güçlü striatal aktivasyonun DRD2 genindeki DNA metilasyonu ile bağlantılı olduğunu göstermiştir. Ek olarak, DRD2 metilasyonu alkol kullanım bozukluğu (AUD) şiddeti ile bağlantılıydı. Özellikle, DRD2 metilasyonu AUD Tanımlama Testi, Bozuk Kontrol Ölçeği ve Alkol Bağımlılığı Ölçeği’ndeki puanlarla bağlantılıydı.
Alkol ve çeşitli diğer yasadışı uyuşturucuların kronik tüketimi, yönetici kontrolle bağlantılı sinir ağları içindeki işlevsel bağlantı eksiklikleri de dahil olmak üzere olumsuz sonuçlarla ilişkilendirilmiştir. 113 Bu eksiklikler, sapkın alkol arama davranışının gelişimine dolaylı olarak katkıda bulunabilir. 113 Hagerty ve ark . 114 özellikle DRD2 genindeki ortalama DNA metilasyonunun sol ve sağ yönetici kontrol ağı bağlantısıyla negatif korelasyonlu olduğunu, ancak test edilen diğer ağlarla hiçbir korelasyon bulunmadığını bulmuştur. Ek olarak, DRD2 DNA metilasyonunun alkol sorunlarının ciddiyetiyle bağlantılı olduğu bulunmuştur. Bu bulgular, çoklu madde kullanıcıları arasında alkol tüketiminin nörobiyolojik belirteçlerini epigenetik etkilere bağlayan teorik bir çerçeveyi desteklemektedir.
AUD’li bireylerde dopaminerjik değişiklik görülme eğilimi vardır ve ailelerinde AUD öyküsü olanlar beyin gelişiminde etkiler yaşayabilirler. 115 Hill ve Sharma, ailesel yüksek risk durumu ile DRD2 genindeki DNA metilasyonu arasında anlamlı bir pozitif korelasyon keşfettiler. 116 Aslında, yüksek riskli ve düşük riskli ailesel risk grupları arasında sol inferior temporal, sol fusiform ve sol insula bölgelerinin hacminde anlamlı farklılıklar gözlemlendi. Daha önce, bu bölgeler sosyal bilişle ilişkilendirilmişti. Ek olarak, DRD2 geninin DNA metilasyonu bu bölgelerdeki gri madde hacimleriyle ters orantılıydı.
Benzer düşünce çizgisinde, Hillemacher ve ark ., 117 epigenetiğin dopaminerjik nörotransmisyondaki rolü ve alkol bağımlılığı üzerindeki etkisiyle ilgili olarak, alkol yoksunluğu/erken yoksunluk sırasında DRD2 geninde DNA metilasyonunda önemli bir artış buldular. Dahası, obsesif-kompulsif içme ölçeğiyle ölçülen istek ile DRD2 geninin DNA metilasyonu arasında önemli bir ilişki keşfettiler. Alkol isteğiyle ilgili bu ilişkiye dair bulguları, DRD2 geninin DNA metilasyonunun bağımlılık davranışının nörobiyolojisindeki temel rolünü vurgular. Kontroller ve hastalar arasında DNA metilasyonunda önemli değişiklikler gösterilememesinin, kontrol grubunun yetersiz taranmasından kaynaklanabileceğini belirtmekte fayda var. 118
Ayrıca, daha önce birkaç araştırmacı tarafından da belirtildiği gibi, 89 – 104 DRD2’nin ligand bağlanmasının azalması, AUD ve diğer RDS davranışları olan bireylerin striatumunda sürekli olarak gözlemlenmiştir. Azalmış DRD2 bağlanmasının, DRD2 yoğunluğunun azaldığını gösterdiği ve bunun da istekleri tetiklediği ve nüksetme olasılığını artırdığı ileri sürülmüştür. Buna göre, Feltmann ve ark ., 119 şaşırtıcı bir şekilde, diğerlerinden farklı olarak, DRD2 geninin araştırılan bölgelerinde DNA metilasyon farklılıkları bulamadı . Bununla birlikte, Wistar sıçanlarında, kronik alkol tüketimi , NAc’deki DRD2 geninin uzun izoformunun mRNA seviyelerini önemli ölçüde azalttı . Ek olarak, alkol tüketimi DRD2-DRD2 homoreseptör komplekslerinin striatal yoğunluğunu azaltmış, NAc kabuğunda ve dorsal striatumda A2AR-DRD2 heteroreseptör komplekslerinin yoğunluğunu artırmış ve dorsal striatumda sigma1R- DRD2 heteroreseptör komplekslerinin yoğunluğunu azaltmıştır. İlginç bir şekilde, bu sıçanlardaki kronik alkol tüketimi, Blum’un laboratuvarında Altın Suriye hamsterlarını içeren önceki bulgularla iyi örtüşmektedir. 120 Özellikle, sadece su içen kontrol hamsterlarıyla karşılaştırıldığında, bir yıl sonra serbestçe etanol içen deney hamsterlarının bazal ganglionlarında belirgin şekilde daha düşük bir lösin-enkefalin benzeri immünoreaktif madde konsantrasyonu vardı. 120
Bu keşif, etanolün etkilerinin, Feltmann ve ark . çalışmasında sunulan uzun süreli içki içme bulgularına benzeyen endojen peptidil opiatların sentezini içerdiğini ileri sürmektedir . Bu nedenle, diğer bölgelerde DNA metilasyon farklılıklarının ve/veya Feltmann ve ark . 119 çalışmasında incelenmeyen diğer epigenetik modifikasyonların meydana gelme olasılığını göz ardı edemesek de, alkol içerek mRNA ekspresyonunun baskılanmasının DRD2 sentezini de azaltma olasılığı sorusunu gündeme getirmektedir. Feltmann ve ark . 119 ayrıca AUD’nin hipodopaminerjik bir sistemle ilişkili olduğu hipotezini desteklemiş ve A2AR-DRD2 heteroreseptör kompleksini tedavi için umut vadeden yeni bir hedef olarak önermiştir.
5.1.4. Psikostimülanların kötüye kullanımı
Son zamanlarda, Blum ve diğerleri 30 belirli terapötik modalitelerle elde edilen hafif D2 reseptör uyarımının dopamin salınımına neden olabileceğini ve bunun da D2 yönlendirmeli mRNA’yı değiştirebileceğini ve insanlarda DRD2 işlevini iyileştirebileceğini ileri sürmüşlerdir. DRD2 aktivitesindeki bu artışın, özellikle yüksek riskli, genetik olarak zayıflamış popülasyonlarda, istek davranışlarını azalttığı düşünülmektedir. Buna göre, Cadet’in grubu, epigenetik zararlar açısından metamfetamin (METH) ve modafinili karşılaştıran etkili deneyler yürütmüştür. 121
González ve ark ., 121 farelere METH veya modafinilin tekrarlanan uygulanmasının bilişsel etkilere yol açtığını göstermiştir. Medial prefrontal kortekste, histon asetilasyonu ve DNA metilasyon profillerinde indüksiyonlar gözlemlenmiştir. Tekrarlanan METH maruziyetine maruz bırakılan fareler (modafinile maruz bırakılmayanlar hariç), yeni nesne tanıma testi ile ortaya konduğu gibi, hafıza tanımada belirgin bir azalma ile bozulmuş bilişsel hafıza sergilemiştir. Ek olarak, METH ile tedavi edilen fareler (i) histon H3 ve H4 asetilasyonu seviyelerinde azalma ve 5-mC seviyelerinde artış ve (ii) DRD2 geninin promotörlerinde azalmış histon H3 asetilasyonu zenginleşmesi göstermiştir. Bu bulgular, özellikle DRD2 genindeki epigenetik düzensizliğin, METH’nin uzun vadeli bilişsel gerileme etkileri ve medial prefrontal korteks fonksiyonu üzerindeki olumsuz etkileri ile ilişkili olduğunu düşündürmektedir. Psikostimülan kötüye kullanımından etkilenen epigenetik düzenlemenin spesifik mekanizmalarını açıklamak için daha fazla çalışmaya ihtiyaç vardır.
NAc ve lizin kalıntıları içindeki DNA’daki histon metilasyonu ve asetilasyonundaki değişiklikler tekrarlanan kokain uygulamasıyla gözlemlenmiştir. Nestler’in grubu, ödül işlemeyle ilgili modellerde histon arginin (R) metilasyonunu araştırmıştır. Özellikle, Damez-Werno ve ark . 122 hem hayvan hem de insan kendi kendine uygulama deneylerinde, histon işaret proteini-R-metiltransferaz-6 (PRMT6) ve histon H3’teki R2’nin asimetrik demetilasyonunun (H3R2me2a) kemirgen ve kokain bağımlı insan NAc’de azaldığını bulmuştur. Aslında, DRD1 (D1-MSN’ler) ifade eden orta dikenli nöronlarda (MSN’ler) PRMT6 aşırı ekspresyonu kokain arama davranışlarına karşı koruyucu olsa da, tüm NAc nöronlarındaki D2-MSN’lerde PRMT6 aşırı ekspresyonu artan kokain arama davranışlarıyla ilişkilendirilmiştir. Bu doğrultuda, Blum ve ark . PUD’de dopaminilasyonun (H3R2me2a bağlanması) meydana geldiği ve bağlayıcı inhibitör Srcin1’in, majör DRD2 A2 alelik polimorfizmi gibi, NAc dopamin ekspresyonunu normalleştirerek psikostimülan arama davranışına karşı koruduğu hipotezi ileri sürülmüştür. Dahası, çok sayıda çalışma DRD2 Taq A1 aleli (30 – 40 daha düşük DRD2 sayısı) ile şiddetli kokain bağımlılığı arasındaki ilişkiyi doğrulamıştır. Lepack ve diğerlerine göre , 123 akut kokain, NAc sinapslarındaki dopamini artırır ve histon H3 glutamin 5 dopaminilasyonuna ve ardından DRD2 ekspresyonunun inhibisyonuna neden olur. Uzun süreli kokain kullanımıyla, DRD2 ekspresyonunun inhibisyonu artar ve kokain yoksunluğuna eşlik eder. Dahası, kokain yoksunluğu sırasında Src kinaz sinyal inhibitörü 1’in (Srcin1 veya p140CAP) H3R2me2a bağlanmasını azalttığını bildirmişlerdir. Dolayısıyla, bu inhibe edilen dopaminilasyon bir “homeostatik fren” indüklemiştir. Blum’un 37. grubu, DRD2 Taq A2 aleli gibi NAc D2-MSN’lerde Src sinyallemesindeki azalmanın , SUD’ye karşı koruyucu iyi bilinen bir genetik mekanizma, NAc dopamin ekspresyonunu normalleştirdiğini ve kokain isteklerini ve kokain arama davranışlarını azalttığını ileri sürmüştür. Bu nedenle, Srcin1 terapötik müdahaleler için önemli bir hedef olabilir.
METH kaynaklı psikozla ilgili bir diğer önemli yön, hem genetik DNA öncüllerini hem de örneğin DRD2 genindeki mRNA ekspresyonundaki epigenetik translasyon sonrası değişiklikleri içerebilir. Nohesara ve arkadaşları 124 METH’in METH psikozu olan hastalarda bir dizi dopaminle ilişkili genin (yani, DRD3 , DRD4 ve COMT ) DNA metilasyonunda bir azalmaya neden olduğunu ancak METH olmayan psikoz hastalarında böyle bir azalmaya neden olmadığını buldular. Buradaki öneri, genel olarak METH bağımlılığının DNA metilasyonunda bir azalma ve açık bir kromatin konformasyonu ile bağlantılı olduğu ve bunun psikotik bozuklukların patogenezinde rol oynayan birkaç önemli genin ekspresyonunun artmasına yol açtığıdır. 121 Nohesara ve arkadaşları 124 bu epigenetik değişikliklerin METH bağımlılarında psikoz teşhisi için değerli tanısal biyobelirteçler olarak hizmet edebileceğini öne sürdüler. Ayrıca, bu hastalarda psikozun baskılanması veya önlenmesi için metil açısından zengin bir diyetin kullanımını desteklemekte ve böylece daha geniş popülasyonları içeren daha fazla ilişki ve müdahale çalışmasını teşvik etmektedirler.
5.1.5. Opioid kötüye kullanımı
Opioid kullanım bozukluğu (OUD) ve diğer ödül-düzensiz bozuklukların kalıtım derecesi yüksektir. Zhang ve ark . 125 DRD1 ve DRD2 genlerindeki çeşitli metilasyon niceliksel özellik lokuslarının (mQTL’ler) hem sağlıklı kontrol hem de eroin kullanım bozukluğu gruplarında tanımlandığını göstermiştir. Özellikle, DRD1 genindeki rs4867798-CpG_174872884 ve rs5326-CpG_174872884, eroin kullanım bozukluğu olan hastalarda gözlemlenen benzersiz tek nükleotid polimorfizm-CpG çiftleriydi. Bu çığır açan araştırma, belirli dopaminerjik mQTL’lerin DNA metilasyonu ve gen ekspresyonunu ima ederek OUD özelliklerine bağlanabileceğini öne sürmektedir. Ancak, DRD2 ile ilgili bulguları yeniden değerlendirmek için kontrollerin daha iyi taranmasının gerekliliğini vurgulamaktadır. 120
5.1.6. Yeme bozuklukları
SUD’a benzer şekilde, anoreksiya nervoza çok yönlü ve oldukça kalıtımsal bir hastalıktır. 126 – 128 Rask-Andersen ve arkadaşları 128 ilgili literatürü gözden geçirdiler ve 43 genle ilişkili 128 farklı polimorfizmi inceleyen bir anoreksiya nervoza kohortunda 175 ilişki çalışması yürütüldüğünü keşfettiler. En güçlü korelasyonlar, belirli dopaminerjik genlerin vücut kitle indeksini düzenlemede önemli bir rol oynadığını göstermektedir. 129 Dahası, Frieling ve arkadaşlarının bulguları 130 dopamin taşıyıcısı ( DAT ) mRNA ekspresyonunda bir artış ve DRD2 ekspresyonunda bir azalma olduğunu ortaya koymuştur. Frieling ve arkadaşları , DAT geninin yukarı regülasyonunun anoreksiya nervoza ve bulimia nervoza grubunda genin promotörünün hipermetilasyonuyla birlikte olduğunu, DRD2 promotörünün önemli hipermetilasyonunun ise yalnızca anoreksiya nervoza grubunda mevcut olduğunu öne sürmüşlerdir. İncelenen diğer çeşitli dopamin reseptörlerinde metilasyon veya ifadede herhangi bir tutarsızlık tespit edilmedi. Ek olarak, Groleau ve ark . 131 bulimia spektrum bozukluğu tanısı konan kadınların, özellikle borderline kişilik bozukluğu olanların, yeme bozukluğu olmayan kadınlara kıyasla DRD2 DNA metilasyon seviyelerinde önemli yükselmeler gösterdiğini buldu. Dahası, bulimia spektrum bozukluğu ve çocukluk çağı cinsel istismarı öyküsü olan kadınlar, yeme bozukluğu öyküsü olmayan gruba kıyasla artmış DRD2 DNA metilasyonu gösterdi. Bu sonuçlar, bulimia spektrum bozukluğu olan bireylerin DRD2 gen promotörünün DNA metilasyonunda bir artış yaşadığını ve bunun genel olarak yeme bozukluklarının bir kurulu korelasyonu olmaktan ziyade komorbid psikopatolojinin daha güçlü bir belirteci olarak hizmet edebileceğini göstermektedir.
5.1.7. Kumar
Çeşitli çalışmalara göre, dopaminerjik devreler sorunlu kumar oynama davranışının patofizyolojisinde rol oynayabilir. 132 – 134 Hillemacher ve ark . 134 DRD2 genindeki DNA metilasyon modellerinin 12 aylık veya 30 aylık bir süre boyunca yoksunluk açısından değiştiğini bildirmiştir. Ayrıca, yoksunluk yaşamayan ve tedavi aramayan bireylerde DNA metilasyon seviyelerinde artış gözlemlemişlerdir. Bu bulgular, DNA metilasyonundaki değişikliklerden kaynaklanan değişmiş DRD2 ekspresyonunun, yaşam boyu patolojik kumar oynama davranışında patofizyolojik öneme sahip olduğunu düşündürmektedir.
5.1.8. Kişilik bozuklukları
Kişilik özelliklerinin beş boyutunu (uyumluluk, baskınlık, dışa dönüklük, açıklık ve tepkisellik/güvenilmezlik) sergilediği bilinen şempanzeleri kullanan Staes vd . 135 , DRD2 DNA metilasyonunun dışa dönüklükle en güçlü şekilde bağlantılı olduğunu gözlemlediler. Ek olarak, belirli DRD2 bölgelerindeki değişen DNA metilasyon seviyelerinin , anaokulunda yetiştirilen ancak anne tarafından yetiştirilmeyen bireylerde dışa dönüklükteki değişikliklerle bağlantılı olduğunu buldular. Bu bulgular, erken yaşamda biyolojik anne ile ilişkili yetiştirmenin önemine dair daha fazla destek sağlıyor. Bu çalışma, erken yaşam deneyimlerinin, teorik olarak epigenetik modifikasyon yoluyla uzun süreli davranışsal etkileri etkileyebileceğini ileri sürüyor. Buna göre, bu sonuçlar, kişilik gelişiminde deneyime bağlı metilomun kritik rolünü gösteren artan araştırma gövdesine katkıda bulunuyor. 134 , 136 – 140
5.1.9. Anne yoksunluğu (MD)
Gondre-Lewis’in grubu, yaşamın erken dönemlerinde yaşanan stresin, ergenlerin ve yetişkinlerin yaşamlarının ilerleyen dönemlerinde duygusal ve bağımlılık davranışlarını etkileyen karmaşık bir nörokimyasal kaskadı tetikleyebileceğini doğru bir şekilde belirtmektedir. 141 Dahası, araştırmalar uyuşturucu arama davranışının ve aşırı alkol tüketiminin sıklıkla depresif benzeri semptomlar ve davranışlarla birlikte bulunduğunu göstermektedir. 142 Bu davranışlar, yaşamlarının erken dönemlerinde sıkıntıyla karşılaşan bireylerde sıklıkla gözlenmektedir ve erken yaşam stresine maruz kalmanın hayvan modellerinde bulunan önemli özelliklerdir. 143 Aslında, Guo vd . 144 sıçanlarda, MD, kronik öngörülemeyen stres (CUS) ve her ikisinin bir kombinasyonuna (MD/CUS) maruz bırakılan sıçanların, normal kontrollerle karşılaştırıldığında artmış seviyelerde DRD2 promotör DNA metilasyonu gösterdiğini bildirmiştir. Yazarlar, erken yaşam MD’sinin yetişkin sıçanlarda stres kaynaklı depresif benzeri davranışlara karşı duyarlılığı artırdığı sonucuna varmışlardır. Gondre-Lewis grubunun bulgularıyla uyumlu olarak, 141-143 ventral tegmental alandaki DRD2 promotör geninin metilasyonunun artması depresyon ve alkol arama davranışı riskini artırabilir. Ayrıca, Zhu vd . 145 MD’nin yetişkin sıçanların mekansal öğrenme ve hafızası, keşifsel ve limbik aktivitesi üzerindeki etkilerini, bunların NAc’deki DAT , DRD1 , DRD2 ve DRD3 ekspresyonuyla ilişkisini araştırdı . Ek olarak, Zhu vd . 145 DRD2 gen ekspresyonunun düzenlenmesinde DNA metilasyonunun potansiyel katılımını araştırdı . Çalışmalarına göre, yalnızca DRD2 mRNA seviyesi toplam mesafe ile ilişkiliydi. Ancak, DNMT1 ve 3 alfa (DNMT3A) ekspresyonu ve DRD2 geninin promotör bölgesindeki metillenmiş CpG seviyeleri MD grubunda kontrol grubuna kıyasla önemli ölçüde değişmedi. 120 Çalışmaya dahil edilmeyen DRD2 genlerinde diğer bölgelerde işlevsel farklılıklar meydana gelmiş olabilir ve/veya diğer epigenetik modifikasyonlar DRD2 mRNA ekspresyonuyla ilişkilendirilmiştir. Bununla birlikte, eksik kontrol taraması ve yorumlamasının ilişki çalışmasını etkilemiş olması mümkündür ve sadece MD değil, dikkatlice seçilmiş kontrollerle daha fazla araştırma yapılması bu sonuçları değiştirebilir.
5.1.10. Sosyal yenilgi stresi
Sosyal yenilgi stresi açısından, Zhu vd . kronik sosyal yenilgi stresinin NAc’de FOSB ekspresyonunu düzenlediğini ve bunun da bu bölgedeki iki MSN alt tipinde hücre tipi-spesifik ΔFosB birikimini desteklediğini bildirdi. ΔFosB, dayanıklı farelerin NAc’sindeki D1-MSN’lerde ve duyarlı farelerin D2-MSN’lerinde seçici olarak indüklenir. 146 Bu bağlamda, Hamilton vd . D2-MSN’lerde FOSB hedefli histon asetilasyonu veya D1-MSN’lerde histon metilasyonunun strese duyarlı, depresif benzeri bir fenotipi desteklediğini, D2-MSN’lerde histon metilasyonunun veya D1-MSN’lerde histon asetilasyonunun ise sosyal etkileşim davranışı ve sakaroz tercihiyle ölçülen sosyal strese karşı dayanıklılığı artırdığını bildirdi. 147 Önemlisi, çalışma, sosyal yenilgi stresi davranışını düzenleyen gerçek dünya transkripsiyonel süreçlerini simüle ederek, histon modifikasyonlarının özellikle genlere ve hücrelere hedeflendiğine dair ilk kanıtı sundu.
6. Etkileyici kısımlar
Etkileyici gruplar, proteinlere seçici olarak bağlanarak biyolojik aktivitelerini değiştiren moleküllerdir. Esasen ligandlar olan bu moleküller, enzim aktivitesini, genomik aktiviteyi, hücre sinyallemesini ve diğer protein fonksiyonlarını değiştirebilir. 35 Kromozom histonlarında epigenetik olarak indüklenen transkripsiyon sonrası metilasyonu veya asetilasyonu belirlemek çok önemli olsa da, transkripsiyonel aktivatör alanlarını düzenlemede ilk ve en başarılı etkili gruplardan biri, herpes simpleks virüsünden türetilen VP64’tü. VP64, gen hedefli transkripsiyonel aktivasyon için doğrudan RNA polimeraz ll’yi işe alır. 148 Artık, VP64’ü ZFP, 149 TALE, 150 , 151 veya CRISPR/dCas9 gibi DNA bağlayıcı alanlar kullanarak tek bir genin promotör bölgesine hedefleyerek, laboratuvarların hedeflenen genlerin transkripsiyonunu in vitro ve hatta in vivo olarak tutarlı bir şekilde indükleyebileceği anlaşılmıştır . 153 , 154
Ayrıca, gen ifadesi üzerinde çift yönlü kontrol elde etmek için, endojen genlerin baskılanması tipik olarak KRAB efektör grupları kullanılarak gerçekleştirilir. KRAB, insan çinko parmak transkripsiyon faktörlerinde bulunan bir transkripsiyonel baskılama alanını temsil eder. Daha sonra H3K9me3 baskılayıcı işaretlerini biriktiren heterokromatin oluşturan kompleksleri işe alarak işlev görür ve bu da transkripsiyonel baskılamaya neden olur. 155 VP64’e benzer şekilde, KRAB alanı enzimatik aktiviteyi kullanmak yerine ikincil faktörleri görevlendirerek epigenomu değiştirir. Ek olarak, promotör veya güçlendirici hedefli KRAB hücre kültüründe ve beyinde kullanılmıştır. 156 , 157 Ek olarak, araştırmalar bağımlılık patogenezi ile ilişkili spesifik transkripsiyon faktörlerini ve epigenetik okuyucuları, silgileri ve yazıcıları tanımlamıştır. Daha yakın zamanda, nöroepigenetik düzenleme için bir dizi efektör grubu geliştirilmiştir. Bu efektör grupları arasında DNMT3A (CpG metilasyonundan sorumludur) gibi DNA’yı değiştiren enzimler bulunur.158 , 159 ve TET1 (CpG’nin hidroksimetilasyonunu katalize eder).160 , 161 Ayrıca , çekirdek histonları asetile etmek için HAT’leri işe alan NFκB alt birimi p65 gibi histon PTM’lerini düzenlemede rol oynayan proteinleri de kapsarlar. 162 histon metiltransferaz G9a (H3K9me2’yi katalize eder), 162 , 163 p300 HAT (dört çekirdek histonu da asetile eder), 164 Sin3 etkileşim alanı (HDAC’leri toplar), 165 , 166 lizin-spesifik demetilaz 1 (H3K4 ve H3K9’u demetile eder), 167 PRDM9 (H3K4 ve H3K36’yı metile eder), 168 ve DOT1L (H3K79’u metile eder). 169 , 170 Ayrıca, CREB gibi transkripsiyon faktörleri de kullanılmış ve böylece bilinen transkripsiyonel düzenleme mekanizmaları taklit edilmiştir. 17
7. SUD epigenetiğinde araştırma görünümü
Bu yorumda sunulan kanıtlar, epigenetik (yanlış) düzenlemenin SUD’deki önemli rolünü güçlü bir şekilde göstermektedir. Çok sayıda ilişki ve müdahaleli çalışma, epigenetik mekanizmaların, özellikle DRD2 geninin ifadesiyle ilgili olarak, dopamin homeostazının karmaşık düzenlenmesinden sorumlu olduğunu göstermektedir . Epigenetik düzenleyici mekanizmalar (yani, histon modifikasyonları, ncRNA ve DNA metilasyonu), genetik arka plandan (örneğin, mutasyonların varlığı) ve çevresel faktörlerden (örneğin, ilaçlardan, alkolden ve tütünden gelen aktif bileşiklerin varlığı) etkilenen fenotipleri oluşturmak için düzenlenmiş bir şekilde etkileşime girer. Daha da önemlisi, bu etkileşimler doku ve hücreye özgü bir şekilde gerçekleşir. Dolayısıyla, SUD’nin çeşitli sunumları boyunca epigenetik modifikasyonların ayrıntılı bir haritalanması, her bir vakada yer alan kesin düzenleyici mekanizmaların anlaşılmasını ve terapötik müdahaleler için potansiyel hedeflerin belirlenmesini sağlayacaktır.
Etkileyici kısımların kullanımı umut verici bir müdahale olarak ortaya çıkmaktadır. Ancak, epigenetik değiştiricilerin (örn. DNMT’ler, HDAC’ler ve HTA’lar) pleyotropik etkileri nedeniyle dikkatli olunmalıdır ve bu enzimlerin sistemik inaktivasyonu istenmeyen yan etkilere yol açarak klinik çevirilerini engelleyebilir. Etkileyici kısımların hücreye özgü aktivasyonu veya iletimi için mekanizmaları açıklamak için daha fazla araştırmaya ihtiyaç vardır. Ayrıca, doğum öncesi SUD’nin yavru sağlığını etkileyebileceği göz önüne alındığında, mekanistik ve müdahaleci çalışmalar epigenetik değişiklikleri belirlemek ve hedeflemek, doğum öncesi SUD’nin fetal sağlık üzerindeki etkilerini azaltmayı amaçlamaktadır.
8. Özet
SUD, genellikle tekrarlanan psikoaktif uyuşturuculara maruz kalmanın ardından duyarlı bireylerde gözlenen kalıcı davranışsal anormallikleri içerir. Bu davranışsal değişikliklerin kalıcı doğası, bağımlılık fenotipine katkıda bulunabilecek belirli beyin bölgelerindeki gen ifadesinde olası uzun süreli değişiklikler olduğunu göstermektedir. Son on yılda yapılan ileri düzey araştırmalar, özellikle beyinde olmak üzere çeşitli dokularda gen ifadesinde pozitif veya negatif olsun, kalıcı değişiklikleri düzenlemede epigenetik mekanizmaların temel katılımını ortaya koymuştur. Bu anlayış, psikoaktif uyuşturucuların beyin üzerindeki kalıcı etkilerinin aracılık etmesinde epigenetik düzenleyici süreçlerin etkisini açıklığa kavuşturmayı amaçlayan araştırmaları teşvik etmiştir; bu araştırmalar, ağırlıklı olarak uyuşturucu bağımlılığının hayvan modellerini kullanmıştır. Ancak, daha yakın zamanda, insan nöroepigenetik araştırmaları hızla ortaya çıkmaktadır. İkna edici kanıtlar, tekrarlanan uyuşturuculara maruz kalmanın, üç temel epigenetik düzenleme modu aracılığıyla beynin ödül bölgelerinde değişikliklere neden olduğunu göstermektedir: Asetilasyon ve metilasyon gibi histon modifikasyonları, DNA metilasyonu ve ncRNA’lar. Bu yorumda, odak noktamız DRD2 genindeki epigenetik değişiklikleri araştırmak ve bu epigenetik değişikliklerin bağımlılıkla ilişkili davranışsal anormalliklerdeki rolünü doğrudan göstermektir. Şu anda, yakın zamanda Pandey’in 104. grubunun alkol kaynaklı kaygıyı hafifletmede gözlemlediği gibi, bu istenmeyen negatif epigenetik değişiklikleri etkisiz hale getirmek için çeşitli efektör kısımlarının kullanımıyla ilgili artan bir farkındalık vardır.
Deneysel olarak, efektör grupları yalnızca mekanik içgörüler için değil, aynı zamanda bu olumsuz epigenetik hakaretleri ortadan kaldırmanın önemini göstermek için de benzersiz bilgiler sağlıyor ve bu da DRD2 geni gibi belirli mRNA transkripsiyonel ifadesinin eş zamanlı zayıflamasına yol açıyor. Cevaplanmamış önemli bir soru, örneğin dopamin homeostazının indüksiyonunun, transkripsiyon sonrası histon metilasyonunun asetilasyona dönüşümünü artırıp artırmayacağıdır. Laboratuvarımız, potansiyel olarak hem hayvan modelleri hem de insanlarda pro-dopamin düzenlemesini nazikçe indüklemek için bir nutrasötik kompleks geliştiriyor. Bu son derece önemli soruyu cevaplamak için, bilim camiasının bu alanda gerekli araştırmayı yapmasını savunuyoruz.
9. Sonuç
DRD2 geni çeşitli nöropsikiyatrik bozukluklarda kapsamlı bir şekilde araştırılmıştır. DRD2 Taq A1 alelinin 1990 yılında şiddetli alkolizmle ilk kez ilişkilendirilmesinin ardından çok sayıda uluslararası çalışma yapılmıştır. Bizim görüşümüze göre, çeşitli DRD2 gen polimorfizmlerinin ilişkisine ilişkin olumsuz raporların birincil nedeni, birçok gizli RDS davranışını ortadan kaldıramayan kontrollerin yetersiz taranmasıdır. Dahası, DRD2 varyantlarının pleiotropik etkileri, bu yorumda tartışıldığı gibi, epigenetik DNA metilasyonu ve negatif histon translasyon sonrası metilasyonunun tanımlandığı nörofizyolojik, nöropsikolojik, stres tepkisi, sosyal stres yenilgisi, MD ve kumar bozukluğu bağlamlarında gözlemlenmiştir. 19 Ekim 2022 itibarıyla, DNA metilasyonuna odaklanan 70 makale ve histon metilasyonuyla ilgili 20 makale PUBMED’de listelenmiştir. Önemlisi, Blum ve Noble, DRD2 Taq A1 alelini alkolizme özgü olarak değil, genelleştirilmiş bir ödül geni olarak tanımladılar. Bu nedenle, şimdi alanın nöroepigenetik hakaretleri düzenlemek için efektör kısımlarını kullanmanın yollarını bulması veya dopamin homeostazını indükleyerek negatif mRNA azaltılmış ifadeyi potansiyel olarak ortadan kaldırma fikrini kullanması gerekiyor. Bu, geleceğe yönelik övgüye değer bir hedefi temsil ediyor.
Teşekkürler
Yazarlar, Margaret A. Madigan’ın uzman düzenlemeleri ve Danielle Kradin’in biçimlendirmesi için teşekkürlerini sunarlar.
Dipnotlar
Çıkar çatışması
Kenneth Blum, pro-dopamin düzenleme kompleksleri ve bağımlılık riski için genetik testlerle ilgili hem yerli hem de yabancı patentlere sahiptir. Diğer yazarlar çıkar çatışması olmadığını beyan etmektedir.
Referanslar
Domi E, Domi A, Adermark L, Heilig M, Augier E. Alkol arama davranışının nörobiyolojisi . J Neurochem . 2021; 157 ( 5 ):1585–1614. doi: 10.1111/jnc.15343 [ PubMed ] [ CrossRef ] [ Google Akademik ]2.
Haass-Koffler CL, Magill M, Cannella N, ve diğerleri. Stres kaynaklı alkol isteği için farmakolojik bir müdahale olarak mifepristone: Bir insan laboratuvar çalışması . Addict Biol . 2023; 28 ( 7 ):e13288. doi: 10.1111/adb.13288 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]3.
Blum K, Chen TJH, Meshkin B, ve diğerleri. Ödül Eksikliği Sendromu’nun (RDS) bir alt türü olan madde arama davranışının azaltılmasını etkilemek için katekol-O-metil-transferaz (COMT) aktivitesinin manipülasyonu gen polimorfizmlerine bağlıdır: Bir hipotez . Med Hipotezleri . 2007; 69 ( 5 ):1054–1060. doi: 10.1016/j.mehy.2006.12.062 [ PubMed ] [ CrossRef ] [ Google Akademik ]4.
Blum K, Trachtenberg MC, Elliott CE, ve diğerleri. Enkefalinaz inhibisyonu ve öncü amino asit yüklemesi alkol ve çoklu uyuşturucu bağımlılarının yatarak tedavisini iyileştiriyor: Besin takviyesi SAAVE’nin çift kör plasebo kontrollü çalışması . Alkol . 1988; 5 ( 6 ):481–493. doi: 10.1016/0741-8329(88)90087-0 [ PubMed ] [ CrossRef ] [ Google Akademik ]5.
Blum K, Chen ALC, Chen TJH, ve diğerleri. Mezolimbik dopaminerjik ödül devresini bloke etmek yerine aktivasyon, ödül eksikliği sendromunun (RDS) uzun vadeli tedavisinde tercih edilen bir yöntemdir: Bir yorum . Theor Biol Med Model . 2008; 5:24 . doi: 10.1186/1742-4682-5-24 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]6.
Blum K, Giordano J, Morse S, ve diğerleri. Büyük zihni anlamak: İnsanlar hala genetik olarak evrimleşiyor . Tne IIOAB . 2010; 1 :1–14. [ Google Akademik ]7.
Comings DE, Blum K. Ödül eksikliği sendromu: Davranış bozukluklarının genetik yönleri . Prog Brain Res . 2000; 126 :325–341. doi: 10.1016/S0079-6123(00)26022-6 [ PubMed ] [ CrossRef ] [ Google Akademik ]10.
Blum K, Chen AL, Oscar-Berman M, ve diğerleri. Ödül eksikliği sendromu (RDS) hastalarında dopaminerjik genlerin nesiller arası ilişki çalışmaları: Ödül bağımlılığı davranışları için uygun fenotiplerin seçilmesi . Int J Environ Res Public Health . 2011; 8 ( 12 ):4425–4459. doi: 10.3390/ijerph8124425 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]11.
Febo M, Blum K, Badgaiyan RD, ve diğerleri. Dopamin homeostazı: Ödül eksikliği sendromunda beyin fonksiyonel bağlantısı . Front Biosci (Landmark Ed) . 2017; 22 ( 4 ):669–691. doi: 10.2741/4509 [ PubMed ] [ CrossRef ] [ Google Akademik ]12.
Gold MS, Blum K, Febo M, ve diğerleri. Ödül eksikliği sendromu (RDS) ve anti-ödül sistemleriyle bağlantılı anhedonideki dopaminin moleküler rolü . Front Biosci (Schol Ed) . 2018; 10 ( 2 ):309–325. doi: 10.2741/s518 [ PubMed ] [ CrossRef ] [ Google Akademik ]13.
Borsook D, Linnman C, Faria V, Strassman AM, Becerra L, Elman I. Ağrı kronifikasyonunda ödül eksikliği ve anti-ödül . Neurosci Biobehav Rev. 2016; 68 :282–297. doi: 10.1016/j.neubiorev.2016.05.033 [ PubMed ] [ CrossRef ] [ Google Akademik ]14.
Bowirrat A, Oscar-Berman M. Dopaminerjik nörotransmisyon, alkolizm ve ödül eksikliği sendromu arasındaki ilişki . Am J Med Genet B Neuropsychiatr Genet . 2005; 132b ( 1 ):29–37. doi: 10.1002/ajmg.b.30080 [ PubMed ] [ CrossRef ] [ Google Akademik ]15.
Filippi A, Mueller T, Driever W. Vglut2 ve gad ekspresyonu, zebra balığı beynindeki dopaminerjik ve noradrenerjik nöronların ikili GABAerjik ve glutamaterjik kotransmitter fenotiplerinin farklı modellerini ortaya koymaktadır . J Comp Neurol . 2014; 522 ( 9 ):2019–2037. doi: 10.1002/cne.23524 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]16.
Valentino RJ, Koroshetz W, Volkow ND. Opioid salgınının nörobiyolojisi: Temel ve çevirisel perspektifler . Biol Psychiatry . 2020; 87 ( 1 ):2–3. doi: 10.1016/j.biopsych.2019.09.003 [ PubMed ] [ CrossRef ] [ Google Akademik ]17.
Browne CJ, Godino A, Salery M, Nestler EJ. Opioid bağımlılığının epigenetik mekanizmaları . Biol Psychiatry . 2020; 87 ( 1 ):22–33. doi: 10.1016/j.biopsych.2019.06.027 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]18.
Rosell DR, Siever LJ. Saldırganlık ve şiddetin nörobiyolojisi . CNS Spectr . 2015; 20 ( 3 ):254–279. doi: 10.1017/S109285291500019X [ PubMed ] [ CrossRef ] [ Google Akademik ]19.
Mahna D, Puri S, Sharma S. DNA metilasyon imzaları: Uyuşturucu ve alkol kötüye kullanımının biyobelirteçleri . Mutat Res Rev Mutat Res . 2018; 777 :19–28. doi: 10.1016/j.mrrev.2018.06.002 [ PubMed ] [ CrossRef ] [ Google Akademik ]20.
D’Aquila PS, Elia D, Galistu A. Susamış ve su yalayan sıçanlarda yutma davranışının aktivasyonunda dopamin D 1 benzeri ve D 2 benzeri reseptörlerin rolü . Psikofarmakoloji (Berl) . 2019; 236 ( 12 ):3497–3512. doi: 10.1007/s00213-019-05317-w [ PubMed ] [ CrossRef ] [ Google Akademik ]21.
Volkow ND, Morales M. İlaçlar üzerindeki beyin: Ödülden bağımlılığa . Hücre . 2015; 162 ( 4 ):712–725. doi: 10.1016/j.cell.2015.07.046 [ PubMed ] [ CrossRef ] [ Google Akademik ]22.
Yamamoto K, Fontaine R, Pasqualini C, Vernier P. Omurgalılarda dopamin reseptör genlerinin sınıflandırılması: Osteichthyes’te dokuz alt tip . Brain Behav Evol . 2015; 86 ( 3–4 ):164–175. doi: 10.1159/000441550 [ PubMed ] [ CrossRef ] [ Google Akademik ]23.
Noble EP, Blum K, Ritchie T, Montgomery A, Sheridan PJ. Alkolizmde D2 dopamin reseptör geninin reseptör bağlama özellikleriyle alelik ilişkisi . Arch Gen Psychiatry . 1991; 48 ( 7 ):648–654. doi: 10.1001/archpsyc.1991.01810310066012 [ PubMed ] [ CrossRef ] [ Google Akademik ]24.
Volkow ND, Chang L, Wang GJ, ve diğerleri. Metamfetamin bağımlılarında beyin dopamin D2 reseptörlerinin düşük seviyesi: Orbitofrontal korteksteki metabolizmayla ilişkisi . Am J Psychiatry . 2001; 158 ( 12 ):2015–2021. doi: 10.1176/appi.ajp.158.12.2015 [ PubMed ] [ CrossRef ] [ Google Akademik ]25.
Volkow ND, Wang GJ, Telang F, ve diğerleri. Düşük dopamin striatal D2 reseptörleri obez kişilerde prefrontal metabolizma ile ilişkilidir: Olası katkıda bulunan faktörler . Neuroimage . 2008; 42 ( 4 ):1537–1543. doi: 10.1016/j.neuroimage.2008.06.002 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]26.
Noble EP, Blum K, Khalsa ME, ve diğerleri. D2 dopamin reseptör geninin kokain bağımlılığı ile alelik ilişkisi . İlaç Alkol Bağımlılığı . 1993; 33 ( 3 ):271–285. doi: 10.1016/0376-8716(93)90113-5 [ PubMed ] [ CrossRef ] [ Google Akademik ]27.
Deng XD, Jiang H, Ma Y, ve diğerleri. DRD2/ANKK1 TaqIA polimorfizmi ile yaygın yasadışı uyuşturucu bağımlılığı arasındaki ilişki: Bir meta-analizden elde edilen kanıtlar . Hum Immunol . 2015; 76 ( 1 ):42–51. doi: 10.1016/j.humimm.2014.12.005 [ PubMed ] [ CrossRef ] [ Google Akademik ]28.
Vereczkei A, Barta C, Magi A, ve diğerleri. Madde kullanımı ve bağımlılık davranışlarının ortak genetik faktörleri olarak FOXN3 ve GDNF polimorfizmleri . J Pers Med . 2022; 12 ( 5 ):690. doi: 10.3390/jpm12050690 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]29.
Dackis CA, Gold MS. Bromokriptin kokain kötüye kullanımının tedavisi olarak . Lancet . 1985; 1 ( 8438 ):1151–1152. doi: 10.1016/s0140-6736(85)92448-1 [ PubMed ] [ CrossRef ] [ Google Akademik ]30.
Blum K, Cadet JL, Gold MS. Metamfetamin ve opioid-dopamin bağlantısını vurgulayan psikostimülan kullanım bozukluğu: Hipodopaminerjik bir çukurdan çıkmak . J Neurol Sci . 2021; 420 :117252. doi: 10.1016/j.jns.2020.117252 [ PubMed ] [ CrossRef ] [ Google Akademik ]31.
Bogomolova EV, Rauschenbach IY, Adonyeva NV, Alekseev AA, Faddeeva NV, Gruntenko NE. Dopamin, Drosophila’da alkalin fosfataz aktivitesini aşağı düzenler : D2 benzeri reseptörlerin rolü . J Insect Physiol . 2010; 56 ( 9 ):1155–1159. doi: 10.1016/j.jinsphys.2010.03.014 [ PubMed ] [ CrossRef ] [ Google Akademik ]32.
Nestler EJ, Peña CJ, Kundakovic M, Mitchell A, Akbarian S. Ruhsal hastalığın epigenetik temeli . Sinirbilimci . 2016; 22 ( 5 ):447–463. doi: 10.1177/1073858415608147 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]33.
Cadet JL, McCoy MT, Jayanthi S. Epigenetik ve bağımlılık . Clin Pharmacol Ther . 2016; 99 ( 5 ):502–511. doi: 10.1002/cpt.345 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]34.
Blum K, Bowirrat A, Gondre Lewis MC, ve diğerleri. Ergen beyin gelişimi sırasında epigenetik durum hiperdopaminerjisinin (Fazlalık) ve genetik özellik hipodopaminerjisinin (Eksik) araştırılması . Curr Psychopharmacol . 2021. doi: 10.2174/2211556010666210215155509 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]50.
Blum K, McLaughlin T, Modestino EJ, ve diğerleri. Arttırılmış beyin ödül fonksiyonel bağlantısı ve dopaminerjik homeostatik sinyallemenin indüksiyonu ile korkunç berrak rüyaların epigenetik onarımı . Curr Psychopharmacol . 2021. doi: 10.2174/2211556010666210215153513 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]51.
Edwards D, Roy AK 3rd, Boyett B, ve diğerleri. Bağımlılık başka bir adla da olsa bağımlılıktır: Ödül eksikliğinin moleküler nörogenetik/epigenetik temellerini benimsemek . J Addict Sci . 2020; 6 ( 1 ):1–4. [ PMC ücretsiz makalesi ] [ PubMed ] [ Google Akademik ]52.
Blum K, Brodie MS, Pandey SC, ve diğerleri. Çoklu uyuşturucu kullanımında alkol aşırı içmenin azaltılmasının araştırılması: Genetik bağımlılık riski şiddeti (GARS) ile birlikte KCNK13 ve RASGRF2 Gen(ler)i risk polimorfizmleri hassas pro-dopamin düzenlemesini yönlendiriyor . J Pers Med . 2022; 12 ( 6 ):1009. doi: 10.3390/jpm12061009 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]53.
Zhang X, Yu H, Bai R, Ma C. Entegre biyoenformatik analiz ile biyobelirteçlerin tanımlanması ve karakterizasyonu ve opioid bağımlılığındaki rolleri . Front Neurosci . 2020; 14 :608349. doi: 10.3389/fnins.2020.608349 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]54.
Law PP, Holland ML. Gen ve çevre etkileşimlerinin kavşağında DNA metilasyonu . Essays Biochem . 2019; 63 ( 6 ):717–726. doi: 10.1042/EBC20190031 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]55.
Moore LD, Le T, Fan G. DNA metilasyonu ve temel işlevi . Nöropsikofarmakoloji . 2013; 38 ( 1 ):23–38. doi: 10.1038/npp.2012.112 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]57.
Jang HS, Shin WJ, Lee JE, Do JT. Epigenetik gen düzenlemesinde ve beyin fonksiyonunda CpG ve CpG Olmayan Metilasyon . Genler (Basel) . 2017; 8 ( 6 ):148. doi: 10.3390/genes8060148 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]58.
Bogdanović O, Lister R. DNA metilasyonu ve hücre kimliğinin korunması . Curr Opin Genet Dev . 2017; 46 :9–14. doi: 10.1016/j.gde.2017.06.007 [ PubMed ] [ CrossRef ] [ Google Akademik ]59.
Sivalingam K, Samikkannu T. Pirasetamın astrositlerde DNA metilasyonunun kokain kaynaklı nöroepigenetik modifikasyonuna karşı nöroprotektif etkisi . Brain Sci . 2020; 10 ( 9 ):611. doi: 10.3390/brainsci10090611 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]60.
Kulis M, Esteller M. DNA metilasyonu ve kanser . Adv Genet . 2010; 70 :27–56. doi: 10.1016/B978-0-12-380866-0.60002-2 [ PubMed ] [ CrossRef ] [ Google Akademik ]61.
Dawson MA, Kouzarides T. Kanser epigenetiği: Mekanizmadan tedaviye . Hücre . 2012; 150 ( 1 ):12–27. doi: 10.1016/j.cell.2012.06.013 [ PubMed ] [ CrossRef ] [ Google Akademik ]62.
Nishiyama A, Nakanishi M. Kanser DNA metilasyon manzarasında gezinme . Trendler Genet . 2021; 37 ( 11 ):1012–1027. doi: 10.1016/j.tig.2021.05.002 [ PubMed ] [ CrossRef ] [ Google Akademik ]63.
Meng H, Cao Y, Qin J, ve diğerleri. DNA metilasyonu, aracıları ve genom bütünlüğü . Int J Biol Sci . 2015; 11 ( 5 ):604–617. doi: 10.7150/ijbs.11218 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]64.
Cavalli G, Heard E. Epigenetikteki ilerlemeler genetiği çevre ve hastalıkla ilişkilendirir . Nature . 2019; 571 ( 7766 ):489–499. doi: 10.1038/s41586-019-1411-0 [ PubMed ] [ CrossRef ] [ Google Akademik ]65.
Cassidy SB, Schwartz S. Prader-Willi ve Angelman sendromları. Genomik baskı bozuklukları . Tıp (Baltimore) . 1998; 77 ( 2 ):140–151. doi: 10.1097/00005792-199803000-00005 [ PubMed ] [ CrossRef ] [ Google Akademik ]67.
Liu C, Jiao C, Wang K, Yuan N. DNA metilasyonu ve psikiyatrik bozukluklar . Prog Mol Biol Transl Sci . 2018; 157 :175–232. doi: 10.1016/bs.pmbts.2018.01.006 [ PubMed ] [ CrossRef ] [ Google Akademik ]68.
Zhang X, Fu R, Yu J, Wu X. DNA demetilasyon: Genetiğin epigenetikle buluştuğu yer . Curr Pharm Des . 2014; 20 ( 11 ):1625–1631. doi: 10.2174/13816128113199990546 [ PubMed ] [ CrossRef ] [ Google Akademik ]69.
Bochtler M, Kolano A, Xu GL. DNA demetilasyon yolları: Ek oyuncular ve düzenleyiciler . Bioessays . 2017; 39 ( 1 ):1–13. doi: 10.1002/bies.201600178 [ PubMed ] [ CrossRef ] [ Google Akademik ]70.
Kafer GR, Li X, Horii T, ve diğerleri. 5-Hidroksimetilsitozin DNA hasarının olduğu yerleri işaretler ve genom stabilitesini destekler . Cell Rep . 2016; 14 ( 6 ):1283–1292. doi: 10.1016/j.celrep.2016.01.035 [ PubMed ] [ CrossRef ] [ Google Akademik ]71.
Chen ZX, Riggs AD. Memelilerde DNA metilasyonu ve demetilasyon . J Biol Chem . 2011; 286 ( 21 ):18347–18353. doi: 10.1074/jbc.R110.205286. [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]72.
Wu X, Zhang Y. TET aracılı aktif DNA demetilasyonu: Mekanizma, işlev ve ötesi . Nat Rev Genet . 2017; 18 ( 9 ):517–534. doi: 10.1038/nrg.2017.33 [ PubMed ] [ CrossRef ] [ Google Akademik ]73.
Ross SE, Bogdanovic O. TET enzimleri, DNA demetilasyonu ve pluripotansiyelliği . Biochem Soc Trans . 2019; 47 ( 3 ):875–885. doi: 10.1042/BST20180606 [ PubMed ] [ CrossRef ] [ Google Akademik ]74.
An J, González-Avalos E, Chawla A, ve diğerleri. TET fonksiyonunun akut kaybı farelerde agresif miyeloid kansere yol açar . Nat Commun . 2015; 6 :10071. doi: 10.1038/ncomms10071 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]75.
Zentner GE, Henikoff S. Histon modifikasyonları ile nükleozom dinamiklerinin düzenlenmesi . Nat Struct Mol Biol . 2013; 20 ( 3 ):259–266. doi: 10.1038/nsmb.2470 [ PubMed ] [ CrossRef ] [ Google Akademik ]77.
Tessarz P, Kouzarides T. Nükleozom yapısını ve dinamiklerini düzenleyen histon çekirdek modifikasyonları . Nat Rev Mol Cell Biol . 2014; 15 ( 11 ):703–708. doi: 10.1038/nrm3890 [ PubMed ] [ CrossRef ] [ Google Akademik ]78.
Stillman B Histon modifikasyonları: Gen ifadesi üzerindeki etkilerine dair içgörüler . Hücre . 2018; 175 ( 1 ):6–9. doi: 10.1016/j.cell.2018.08.032 [ PubMed ] [ CrossRef ] [ Google Akademik ]79.
Hyland EM, Cosgrove MS, Molina H, ve diğerleri. Saccharomyces cerevisiae’deki histon H3 ve histon H4 çekirdek değiştirilebilir kalıntılarının rolüne ilişkin içgörüler . Mol Cell Biol . 2005; 25 ( 22 ):10060–10070. doi: 10.1128/MCB.25.22.10060-10070.2005 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]82.
Varela RB, Resende WR, Dal-Pont GC, ve diğerleri. Amfetamin ve paradoksal uyku yoksunluğu ile indüklenen mani hayvan modellerinde epigenetik düzenleyici enzimlerin rolü . Eur J Neurosci . 2021; 53 ( 2 ):649–662. doi: 10.1111/ejn.14922 [ PubMed ] [ CrossRef ] [ Google Akademik ]83.
Kim S, Kaang BK. Öğrenme ve hafızada epigenetik düzenleme ve kromatin yeniden şekillenmesi . Exp Mol Med . 2017; 49 ( 1 ):e281. doi: 10.1038/emm.2016.140 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]84.
Clapier CR, Cairns BR. Kromatin yeniden şekillendirme komplekslerinin biyolojisi . Annu Rev Biochem . 2009; 78 :273–304. doi: 10.1146/annurev.biochem.77.062706.153223 [ PubMed ] [ CrossRef ] [ Google Akademik ]85.
Han P, Hang CT, Yang J, Chang CP. Kardiyovasküler gelişim ve fizyolojide kromatin yeniden şekillenmesi . Circ Res . 2011; 108 ( 3 ):378–396. doi: 10.1161/CIRCRESAHA.110.224287 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]86.
Beermann J, Piccoli MT, Viereck J, Thum T. Gelişim ve hastalıkta kodlamayan RNA’lar: Arka plan, mekanizmalar ve tedavi yaklaşımları . Physiol Rev . 2016; 96 ( 4 ):1297–1325. doi: 10.1152/physrev.00041.2015 [ PubMed ] [ CrossRef ] [ Google Akademik ]87.
Mattick JS, Makunin IV. Kodlamayan RNA . Hum Mol Genet . 2006; 15 ( Özellik No 1 ):R17–R29. doi: 10.1093/hmg/ddl046 [ PubMed ] [ CrossRef ] [ Google Akademik ]88.
Blum K, Noble EP, Sheridan PJ, ve diğerleri. Alkolizmde insan dopamin D2 reseptör geninin alelik ilişkisi . JAMA . 1990; 263 ( 15 ):2055–2060. [ PubMed ] [ Google Akademik ]89.
Thompson J, Thomas N, Singleton A, ve diğerleri. D2 dopamin reseptör geni (DRD2) Taq1 A polimorfizmi: A1 aleli ile ilişkili insan striatumunda azaltılmış dopamin D2 reseptör bağlanması . Farmakogenetik . 1997; 7 ( 6 ):479–484. doi: 10.1097/00008571-199712000-00006 [ PubMed ] [ CrossRef ] [ Google Akademik ]90.
Savitz J, Hodgkinson CA, Martin-Soelch C, ve diğerleri. DRD2/ANKK1 Taq1A polimorfizmi (rs1800497), sağlıklı kontrollerde ve majör depresif bozukluğu olan hastalarda D2/3 reseptör bağlanması üzerinde zıt etkilere sahiptir . Int J Neuropsychopharmacol . 2013; 16 ( 9 ):2095–2101. doi: 10.1017/S146114571300045X [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]91.
Schellekens AF, Franke B, Ellenbroek B, ve diğerleri. Alkol bağımlılığında ara bir fenotip olarak dopamin reseptör duyarlılığının azalması ve COMT Val158Met ve DRD2 Taq1A genotiplerinin rolü . Arch Gen Psychiatry . 2012; 69 ( 4 ):339–348. doi: 10.1001/archgenpsychiatry.2011.1335 [ PubMed ] [ CrossRef ] [ Google Akademik ]92.
Benton D, Young HA. Beyin dopamin reseptörleri ile obezite arasındaki ilişkinin bir meta-analizi: Yiyecek bağımlılığından ziyade davranış değişiklikleri meselesi mi? Int J Obes (Lond) . 2016; 40 ( Suppl 1 ):S12–S21. doi: 10.1038/ijo.2016.9 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]93.
Eisenstein SA, Bogdan R, Love-Gregory L, ve diğerleri. DRD2/ANKK1 TaqIA alel durumu tarafından striatal D2 reseptör bağlanmasının tahmini . Synapse . 2016; 70 ( 10 ):418–431. doi: 10.1002/syn.21916 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]94.
Shi S, Leites C, He D, ve diğerleri. MikroRNA-9 ve mikroRNA-326 insan dopamin D2 reseptör ekspresyonunu düzenler ve mikroRNA aracılı ekspresyon düzenlemesi genetik bir varyant tarafından değiştirilir . J Biol Chem . 2014; 289 ( 19 ):13434–1344. doi: 10.1074/jbc.M113.535203 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]95.
Völter C, Riedel M, Wöstmann N, ve diğerleri. Sensörimotor kapısı ve D2 reseptör sinyali: Moleküler genetik yaklaşımdan elde edilen kanıtlar . Int J Neuropsychopharmacol . 2012; 15 ( 10 ):1427–1440. doi: 10.1017/S1461145711001787 [ PubMed ] [ CrossRef ] [ Google Akademik ]96.
Sambataro F, Fazio L, Taurisano P, ve diğerleri. Varsayılan mod ağ aktivitesinin ve striatal DAT bağlanmasıyla ilişkisinin DRD2 genotipe dayalı değişimi . Schizophr Bull . 2013; 39 ( 1 ):206–216. doi: 10.1093/schbul/sbr128 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]97.
Frank MJ, Hutchison K. Kaçınma temelli kararlara genetik katkılar: Striatal D2 reseptör polimorfizmleri . Sinirbilim . 2009; 164 ( 1 ):131–140. doi: 10.1016/j.neuroscience.2009.04.048 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]98.
Bertolino A, Fazio L, Di Giorgio A, ve diğerleri. İnsanlarda prefronto-striatal aktivite ve hacim üzerinde dopamin taşıyıcısı ve D2 reseptörü arasındaki genetik olarak belirlenmiş etkileşim . J Neurosci . 2009; 29 ( 4 ):1224–1234. doi: 10.1523/JNEUROSCI.4858-08.2009 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]99.
Bertolino A, Taurisano P, Pisciotta NM, ve diğerleri. Striatal D2 sinyallemesinin genetik olarak belirlenen ölçümleri, çalışma belleği performansı sırasında prefrontal aktiviteyi öngörür . PLoS One . 2010; 5 ( 2 ):e9348. doi: 10.1371/journal.pone.0009348 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]100.
Moyer RA, Wang D, Papp AC, ve diğerleri. İnsan dopamin D2 reseptörünün alternatif eklenmesini etkileyen intronik polimorfizmler kokain kötüye kullanımıyla ilişkilidir . Nöropsikofarmakoloji . 2011; 36 ( 4 ):753–762. doi: 10.1038/npp.2010.208 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]101.
Vercammen A, Weickert CS, Skilleter AJ, Lenroot R, Schofield PR, Weickert TW. Dopaminle ilişkili genlerdeki yaygın polimorfizmler birleşerek ‘şizofreni benzeri’ bir prefrontal hipoaktivite üretir . Transl Psychiatry . 2014; 4 ( 2 ):e356. doi: 10.1038/tp.2013.125 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]102.
Davis C, Levitan RD, Kaplan AS, ve diğerleri. Ödül duyarlılığı ve D2 dopamin reseptör geni: Tıkınırcasına yeme bozukluğunun bir vaka kontrol çalışması . Prog Neuropsychopharmacol Biol Psychiatry . 2008; 32 ( 3 ):620–628. doi: 10.1016/j.pnpbp.2007.09.024 [ PubMed ] [ CrossRef ] [ Google Akademik ]103.
Kraschewski A, Reese J, Anghelescu I, ve diğerleri. Dopamin D2 reseptör geninin alkol bağımlılığıyla ilişkisi: Reseptör işlevini anlamak için anahtar faktörler olarak alkoliklerin haplotipleri ve alt grupları . Pharmacogenet Genomics . 2009; 19 ( 7 ):513–527. [ PubMed ] [ Google Akademik ]104.
Bohnsack JP, Zhang H, Wandling GM, ve diğerleri. Hedeflenen epigenomik düzenleme, ergenlik döneminde alkol maruziyetinden sonra yetişkinlerde anksiyeteyi ve aşırı içmeyi iyileştirir . Sci Adv . 2022; 8 ( 18 ):eabn2748. doi: 10.1126/sciadv.abn2748 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]105.
Oyaci Y, Aytac HM, Pasin O, Çetinay Aydin P, Pehlivan S. Gen varyantları ve klinik parametreler açısından kanabinoid veya sentetik kanabinoid kullanım bozukluğunda MB-COMT promotor ve DRD2 geninin değişmiş metilasyonunun tespiti . J Addict Dis . 2021; 39 ( 4 ):526–536. [ PubMed ] [ Google Akademik ]106.
Gerra MC, Jayanthi S, Manfredini M, ve diğerleri. Esrar kullanımında gen varyantları ve eğitim düzeyi: DNA metilasyonunun aracılık rolü . Transl Psychiatry . 2018; 8 ( 1 ):23. doi: 10.1038/s41398-017-0087-1 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]107.
DiNieri JA, Wang X, Szutorisz H, ve diğerleri. Anne kanabis kullanımı yavrularında ventral striatal dopamin D2 gen düzenlemesini değiştirir . Biol Psychiatry . 2011; 70 ( 8 ):763–769. doi: 10.1016/j.biopsych.2011.06.027 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]108.
Dalterio S, Blum K, DeLallo L, Sweeney C, Briggs A, Bartke A. Farelerde delta 9-THC’ye perinatal maruziyet: Vas deferens’te değişen enkefalin ve norepinefrin duyarlılığı . Subst Alkol Eylemleri Kötüye Kullanımı . 1980; 1 ( 5–6 ):467–471. [ PubMed ] [ Google Akademik ]109.
Liu Q, Xu Y, Mao Y, ve diğerleri. Çinli Han sigara içicilerinde nikotin bağımlılığıyla önemli ölçüde ilişkili NCAM1-TTC12-ANKK1-DRD2 kümesindeki varyantları ortaya çıkaran genetik ve epigenetik analiz . Nikotin Tob Res . 2020; 22 ( 8 ):1301–1309. doi: 10.1093/ntr/ntz240 [ PubMed ] [ CrossRef ] [ Google Akademik ]110.
Roussotte FF, Jahanshad N, Hibar DP, ve diğerleri. Dopamin D2 reseptör geninde (DRD2) uyuşturucu bağımlılığı için bir risk varyantı taşıyıcısı olan yaşlılarda değişmiş bölgesel beyin hacimleri . Beyin Görüntüleme Davranışı . 2015; 9 ( 2 ):213–222. doi: 10.1007/s11682-014-9298-8 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]111.
Pandey SC, Kyzar EJ, Zhang H. Alkol bağımlılığının karanlık tarafının epigenetik temeli . Nörofarmakoloji . 2017; 122 :74–84. doi: 10.1016/j.neuropharm.2017.02.002 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]112.
Bidwell LC, Karoly HC, Thayer RE, ve diğerleri. DRD2 promotör metilasyonu ve alkol ödülünün ölçümleri: Ödül devrelerinin işlevsel aktivasyonu ve klinik şiddet . Addict Biol . 2019; 24 ( 3 ):539–548. doi: 10.1111/adb.12614 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]113.
Morales M, Margolis EB. Ventral tegmental alan: Hücresel heterojenlik, bağlantı ve davranış . Nat Rev Neurosci . 2017; 18 :73–85. doi: 10.1038/nrn.2016.165 [ PubMed ] [ CrossRef ] [ Google Akademik ]114.
Hagerty SL, YorkWilliams SL, Bidwell LC, ve diğerleri. DRD2 metilasyonu, bir grup çoklu madde kullanıcısı arasında yönetici kontrol ağı bağlantısı ve alkol sorunlarının ciddiyetiyle ilişkilidir . Addict Biol . 2020; 25 ( 1 ):e12684. doi: 10.1111/adb.12684 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]115.
Klaus K, Vaht M, Pennington K, Harro J. Popülasyonu temsil eden bir çalışmada DRD2 rs6277 polimorfizminin, çevrenin ve cinsiyetin dürtüsellik üzerindeki etkileşimli etkileri . Behav Brain Res . 2021; 403 :113131. doi: 10.1016/j.bbr.2021.113131 [ PubMed ] [ CrossRef ] [ Google Akademik ]116.
Hill SY, Sharma VK. Alkol bağımlılığı açısından çok yüksek risk taşıyan ailelerden gelen genç yetişkin çocuklarda DRD2 metilasyonu ve bölgesel gri madde hacimleri . Psychiatry Res Neuroimaging . 2019; 286 :31–38. doi: 10.1016/j.pscychresns.2019.03.006 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]117.
Hillemacher T, Rhein M, Burkert A, ve diğerleri. Dopamin reseptörü 2 geninin DNA metilasyonu alkol yoksunluğu sırasında değişir . Eur Neuropsychopharmacol . 2019; 29 ( 11 ):1250–1257. doi: 10.1016/j.euroneuro.2019.09.002 [ PubMed ] [ CrossRef ] [ Google Akademik ]118.
Blum K, Baron D, Lott L, ve diğerleri. Ödül eksikliği sendromu (RDS) içermeyen kontrollerin arayışında: Genetik bağımlılık riski testinde “Kutsal Kase” . Curr Psychopharmacol . 2020; 9 ( 1 ):7–21. [ PMC ücretsiz makale ] [ PubMed ] [ Google Akademik ]119.
Feltmann K, Borroto-Escuela DO, Rüegg J, ve diğerleri. Uzun süreli alkol tüketiminin dopamin D2 reseptörü üzerindeki etkileri: Sıçanlarda striatumdaki gen ifadesi ve heteroreseptör kompleksleri . Alcohol Clin Exp Res . 2018; 42 ( 2 ):338–351. doi: 10.1111/acer.13568 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]120.
Blum K, Briggs AH, Elston SF, ve diğerleri. Uzun süreli etanol maruziyetinden sonra hamster bazal ganglionlarında lösin-enkefalin benzeri immünoreaktif maddenin azalması . Science . 1982; 216 ( 4553 ):1425–1427. doi: 10.1126/science.7089531 [ PubMed ] [ CrossRef ] [ Google Akademik ]121.
González B, Jayanthi S, Gomez N, ve diğerleri. Tekrarlanan metamfetamin ve modafinil, fare medial prefrontal korteksinde farklı bilişsel etkilere ve spesifik histon asetilasyonu ve DNA metilasyon profillerine neden olur . Prog Neuropsychopharmacol Biol Psychiatry . 2018; 82 : 1–11. doi: 10.1016/j.pnpbp.2017.12.009 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]122.
Damez-Werno DM, Sun H, Scobie KN, ve diğerleri. Nucleus accumbens’te kokain etkisinde histon arginin metilasyonu . Proc Natl Acad Sci USA . 2016; 113 ( 34 ):9623–9628. doi: 10.1073/pnas.1605045113 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]123.
Lepack AE, Werner CT, Stewart AF, ve diğerleri. Ventral tegmental alanda histon H3’ün dopaminilasyonu kokain arayışını düzenler . Science . 2020; 368 ( 6487 ):197–201. doi: 10.1126/science.aaw8806 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]124.
Nohesara S, Ghadirivasfi M, Barati M, ve diğerleri. Metamfetamin kaynaklı psikoz, DNA hipometilasyonu ve AKT1 ve anahtar dopaminerjik genlerin artmış ekspresyonu ile ilişkilidir . Am J Med Genet B Neuropsychiatr Genet . 2016; 171 ( 8 ):1180–1189. doi: 10.1002/ajmg.b.32506 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]125.
Zhang J, Fan Y, Zhou J, ve diğerleri. Metilasyon niceliksel özellik lokusu rs5326, eroin kullanım bozukluğu için metadon idame tedavisinin duyarlılığı ve etkili dozajı ile ilişkilidir . Psikofarmakoloji (Berl) . 2021; 238 ( 12 ):3511–3518. doi: 10.1007/s00213-021-05968-8 [ PubMed ] [ CrossRef ] [ Google Akademik ]126.
Munn-Chernoff MA, Johnson EC, Chou YL, ve diğerleri. Yeme bozukluğu ve madde kullanımına bağlı fenotipler arasındaki paylaşılan genetik risk: Genom çapında ilişki çalışmalarından elde edilen kanıtlar . Addict Biol . 2021; 26 ( 1 ):e12880. doi: 10.1111/adb.12880 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]127.
Braun CM, Chouinard MJ. Anoreksiya nervoza nöropsikolojik bir hastalık mıdır? Neuropsychol Rev. 1992; 3 ( 2 ):171–212. doi: 10.1007/BF01108842 [ PubMed ] [ CrossRef ] [ Google Akademik ]128.
Rask-Andersen M, Olszewski PK, Levine AS, ve diğerleri. Anoreksiya nervozanın altında yatan moleküler mekanizmalar: İnsan gen ilişkisi çalışmaları ve gıda alımını kontrol eden sistemlere odaklanma . Brain Res Rev. 2010; 62 ( 2 ):147–164. doi: 10.1016/j.brainresrev.2009.10.007 [ PubMed ] [ CrossRef ] [ Google Akademik ]129.
Wang GJ, Volkow ND, Logan J, ve diğerleri. Beyin dopamini ve obezite . Lancet . 2001; 357 ( 9253 ):354–357. doi: 10.1016/s0140-6736(00)03643-6 [ PubMed ] [ CrossRef ] [ Google Akademik ]130.
Frieling H, Römer KD, Scholz S, ve diğerleri. Yeme bozukluklarında dopaminerjik genlerin epigenetik düzensizliği . Int J Eat Disord . 2010; 43 ( 7 ):577–83. doi: 10.1002/eat.20745 [ PubMed ] [ CrossRef ] [ Google Akademik ]131.
Groleau P, Joober R, Israel M, Zeramdini N, DeGuzman R, Steiger H. Bulimia spektrum bozukluğu olan kadınlarda dopamin D2 reseptörü (DRD2) gen promotörünün metilasyonu: Sınırda kişilik bozukluğu ve çocukluk çağı istismarına maruz kalma ile ilişkiler . J Psychiatr Res . 2014; 48 ( 1 ):121–127. doi: 10.1016/j.jpsychires.2013.10.003 [ PubMed ] [ CrossRef ] [ Google Akademik ]132.
Linnet J Bağımlılıktaki öngörülü dopamin tepkisi: Kumar bozukluğu ve madde kullanım bozukluğunun ortak bir nörobiyolojik temeli mi? Prog Neuropsychopharmacol Biol Psychiatry . 2020; 98 :109802. doi: 10.1016/j.pnpbp.2019.109802 [ PubMed ] [ CrossRef ] [ Google Scholar ]133.
Guerra RF, Batista IR, Kim HS, ve diğerleri. Bozuk kumarbazların striatumunda dopamin taşıyıcı yoğunluğunun nörolojik görüntülemesi . J Gambl Stud . 2023; 39 ( 1 ):119–136. doi: 10.1007/s10899-021-10100-8 [ PubMed ] [ CrossRef ] [ Google Akademik ]134.
Hillemacher T, Frieling H, Buchholz V, ve diğerleri. Dopamin reseptörü 2 geninin DNA metilasyonundaki değişiklikler, yaşam boyu patolojik kumar oynama öyküsü olan bireylerde yoksunluk ve sağlık hizmeti kullanımıyla ilişkilidir . Prog Neuropsychopharmacol Biol Psychiatry . 2015; 63 :30–34. doi: 10.1016/j.pnpbp.2015.05.013 [ PubMed ] [ CrossRef ] [ Google Akademik ]135.
Staes N, White CM, Guevara EE, ve diğerleri. Şempanze Dışadönüklük puanları, dopamin reseptörü geni D2’nin (DRD2) epigenetik modifikasyonu ve erken yetiştirme koşullarına göre değişir . Epigenetik . 2022; 17 ( 12 ):1701–1714. doi: 10.1080/15592294.2022.2058224 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]136.
Juraś-Darowny M, Strzelecki D, Talarowska M. Sınır kişilik – psikanalizden epigenetiğe. Bağlanmanın biyolojik temeli . Psychiatr Pol . 2023:1–15. doi: 10.12740/PP/OnlineFirst/166492 [ PubMed ] [ CrossRef ] [ Google Akademik ]137.
Coelho AA, Lima-Bastos S, Gobira PH, Lisboa SF. Stresle ilişkili bozuklukların nörobiyolojisinde endokannabinoid sinyalizasyonu ve epigenetik modifikasyonlar . Nöronal Sinyal . 2023; 7 ( 2 ):NS20220034. doi: 10.1042/NS20220034 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]138.
Zoratto F, Romano E, Pascale E, ve diğerleri. Kumar oynamaya yatkın bir profil gösteren bireysel sıçanlarda serotonin ve dopamin taşıyıcı genlerinin aşağı düzenlenmesi: Epigenetik mekanizmalar için olası bir rol . Sinirbilim . 2017; 340 :101–116. doi: 10.1016/j.neuroscience.2016.10.041 [ PubMed ] [ CrossRef ] [ Google Akademik ]139.
Cattane N, Rossi R, Lanfredi M, Cattaneo A. Sınırda kişilik bozukluğu ve çocukluk çağı travması: Etkilenen biyolojik sistemleri ve mekanizmaları keşfetme . BMC Psikiyatri . 2017; 17 ( 1 ):221. doi: 10.1186/s12888-017-1383-2 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]140.
McDonald S Bulimia nervoza/bulimia spektrum bozukluğu ve komorbid borderline kişilik bozukluğunun (BN/BSD-BPD) genetiğini ve epigenetiğini anlamak: Sistematik bir inceleme . Eat Weight Disord . 2019; 24 ( 5 ):799–814. doi: 10.1007/s40519-019-00688-7 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]141.
Bassey RB, Gondré-Lewis MC. Kombine erken yaşam stresörleri: Doğum öncesi nikotin ve anne yoksunluğu sıçanlarda duygusal ve uyuşturucu arayan davranışsal fenotipleri etkilemek üzere etkileşime girer . Behav Brain Res . 2019; 359 :814–822. doi: 10.1016/j.bbr.2018.07.022 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]142.
Gondré-Lewis MC, Warnock KT, Wang H, ve diğerleri. Erken yaşam stresi yetişkinlerde aşırı alkol tüketimi ve dürtüsellik için bir risk faktörüdür ve CRF/GABA(A) mekanizması aracılığıyla aracılık edilir . Stres . 2016; 19 ( 2 ):235–247. doi: 10.3109/10253890.2016.1160280 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]143.
Gondré-Lewis MC, Darius PJ, Wang H, Allard JS. Anne yoksunluğu çeken/ayrılmış alkol içen sıçanlarda ödül sistemi çekirdeklerinin stereolojik analizleri . J Chem Neuroanat . 2016; 76 ( Pt B ):122–132. doi: 10.1016/j.jchemneu.2016.02.004 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]144.
Guo Z, Li S, Wu J, Zhu X, Zhang Y. Anne yoksunluğu, ventral tegmental alanda DRD2 promotör metilasyonu yoluyla yetişkin sıçanlarda depresyona karşı duyarlılığı artırdı . Front Psychiatry . 2022; 13 :827667. doi: 10.3389/fpsyt.2022.827667 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]145.
Li T, Peng S, Ma X, Chen X, Zhang X. Yetişkin sıçanlarda anne yoksunluğuna bağlı davranışsal anormallikler, nükleus akumbensteki metilasyon düzenlemesiz D2 reseptör seviyeleriyle ilişkilidir . Behav Brain Res . 2010; 209 ( 2 ):281–288. doi: 10.1016/j.bbr.2010.02.005 [ PubMed ] [ CrossRef ] [ Google Akademik ]146.
Vialou V, Maze I, Renthal W, ve diğerleri. Serum yanıt faktörü, DeltaFosB’nin indüksiyonu yoluyla kronik sosyal strese karşı dayanıklılığı destekler . J Neurosci . 2010; 30 ( 43 ):14585–14592. doi: 10.1523/JNEUROSCI.2496-10.2010 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]147.
Hamilton PJ, Burek DJ, Lombroso SI, ve diğerleri. Fosb genindeki hücre tipi-spesifik epigenetik düzenleme sosyal yenilgi stresine duyarlılığı kontrol eder . Nöropsikofarmakoloji . 2018; 43 ( 2 ):272–284. doi: 10.1038/npp.2017.88 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]148.
Hall DB, Struhl K. VP16 aktivasyon alanı, in vivo protein-protein çapraz bağlanmasıyla belirlendiği gibi çoklu transkripsiyonel bileşenlerle etkileşime girer . J Biol Chem . 2002; 277 ( 48 ):46043–46050. doi: 10.1074/jbc.M208911200 [ PubMed ] [ CrossRef ] [ Google Akademik ]149.
Stege JT, Guan X, Ho T, Beachy RN, Barbas CF 3. Sentetik çinko parmak transkripsiyon faktörleri kullanılarak bitkilerde gen ifadesinin kontrolü . Plant J. 2002; 32 ( 6 ):1077–1086. doi: 10.1046/j.1365-313x.2002.01492.x [ PubMed ] [ CrossRef ] [ Google Akademik ]150.
Crocker J, Stern DL. TALE aracılı transkripsiyonel güçlendiricilerin in vivo modülasyonu . Nat Yöntemleri . 2013; 10 ( 8 ):762–767. doi: 10.1038/nmeth.2543 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]151.
Polstein LR, Perez-Pinera P, Kocak DD, ve diğerleri. TALE ve CRISPR/Cas9 tabanlı transkripsiyonel aktivatörler tarafından DNA bağlanmasının, gen düzenlemesinin ve kromatin yeniden şekillendirilmesinin genom çapında özgüllüğü . Genome Res . 2015; 25 ( 8 ):1158–1169. doi: 10.1101/gr.179044.114 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]152.
Black JB, Adler AF, Wang HG, ve diğerleri. CRISPR/Cas9 tabanlı transkripsiyonel aktivatörler tarafından endojen Lokusların hedeflenen epigenetik yeniden modellenmesi, fibroblastları doğrudan nöronal hücrelere dönüştürür . Hücre Kök Hücresi . 2016; 19 ( 3 ):406–414. doi: 10.1016/j.stem.2016.07.001 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]153.
Zhou H, Liu J, Zhou C, ve diğerleri. CRISPR-dCas9-aktivatör transgenik fareler kullanılarak beyindeki birden fazla genin in vivo eş zamanlı transkripsiyonel aktivasyonu . Nat Neurosci . 2018; 21 ( 3 ):440–446. doi: 10.1038/s41593-017-0060-6 [ PubMed ] [ CrossRef ] [ Google Akademik ]154.
Liao HK, Hatanaka F, Araoka T, ve diğerleri. CRISPR/Cas9 aracılı trans-epigenetik modülasyon yoluyla in vivo hedef gen aktivasyonu . Hücre . 2017; 171 ( 7 ):1495–1507.e15. doi: 10.1016/j.cell.2017.10.025 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]155.
Kim SS, Chen YM, O’Leary E, Witzgall R, Vidal M, Bonventre JV. RING parmak ailesinin yeni bir üyesi olan KRIP-1, çinko parmak proteinlerinin KRAB-A transkripsiyonel baskılayıcı alanıyla ilişkilidir . Proc Natl Acad Sci USA . 1996; 93 ( 26 ):15299–15304. doi: 10.1073/pnas.93.26.15299 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]156.
Groner AC, Meylan S, Ciuffi A, ve diğerleri. KRAB-çinko parmak proteinleri ve KAP1, heterokromatin yayılması yoluyla uzun menzilli transkripsiyonel baskılamayı aracılık edebilir . PLoS Genet . 2010; 6 ( 3 ):e1000869. doi: 10.1371/journal.pgen.1000869 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]157.
Zheng Y, Shen W, Zhang J, ve diğerleri. Beyinde CRISPR interferansına dayalı spesifik ve etkili gen inaktivasyonu . Nat Neurosci . 2018; 21 ( 3 ):447–454. doi: 10.1038/s41593-018-0077-5 [ PubMed ] [ CrossRef ] [ Google Akademik ]158.
Vojta A, Dobrinić P, Tadić V, ve diğerleri. Hedeflenen DNA metilasyonu için CRISPR-Cas9 sisteminin yeniden kullanımı . Nükleik Asitler Res . 2016; 44 ( 12 ):5615–5628. doi: 10.1093/nar/gkw159 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]159.
Stepper P, Kungulovski G, Jurkowska RZ, ve diğerleri. Kimerik dCas9-Dnmt3a-Dnmt3L metiltransferaz ile etkili hedefli DNA metilasyonu . Nükleik Asitler Res . 2017; 45 ( 4 ):1703–1713. doi: 10.1093/nar/gkw1112 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]160.
Liu XS, Wu H, Ji X, ve diğerleri. Memeli genomunda DNA metilasyonunun düzenlenmesi . Hücre . 2016; 167 ( 1 ):233–247.e17. doi: 10.1016/j.cell.2016.08.056 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]161.
Liu XS, Wu H, Krzisch M, ve diğerleri. FMR1 geninin DNA metilasyon düzenlemesiyle kırılgan X sendromu nöronlarının kurtarılması . Hücre . 2018; 172 ( 5 ):979–992.e6. doi: 10.1016/j.cell.2018.01.012 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]162.
Arredondo C, González M, Andrés ME, Gysling K. Akut ve kronik amfetaminin sıçan ventral tegmental alanındaki Nurr1 ve NF-κB p65 üzerindeki zıt etkileri . Brain Res . 2016; 1652 :14–20. doi: 10.1016/j.brainres.2016.09.031 [ PubMed ] [ CrossRef ] [ Google Akademik ]163.
Anderson EM, Sun H, Guzman D, ve diğerleri. Nucleus accumbens kabuğundaki histon di-metiltransferaz G9a’nın devre dışı bırakılması kokain kendi kendine kullanımını, stres kaynaklı yeniden başlamayı ve kaygıyı azaltır . Nöropsikofarmakoloji . 2019; 44 ( 8 ):1370–1376. doi: 10.1038/s41386-018-0305-4 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]164.
Dulman RS, Auta J, Teppen T, Pandey SC. Akut etanol ataksi üretir ve sıçan serebellumundaki histon modifikasyonları yoluyla fmr1 ekspresyonunu indükler . Alcohol Clin Exp Res . 2019; 43 ( 6 ):1191–1198. doi: 10.1111/acer.14044 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]165.
Cong L, Zhou R, Kuo YC, Cunniff M, Zhang F. Doğal TALE DNA bağlayıcı modüllerinin ve transkripsiyonel baskılayıcı alanlarının kapsamlı sorgulanması . Nat Commun . 2012; 3 :968. doi: 10.1038/ncomms1962 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]166.
Kearns NA, Pham H, Tabak B, ve diğerleri. Cas9-histon demetilaz füzyonu ile doğal güçlendiricilerin işlevsel açıklaması . Nat Yöntemleri . 2015; 12 ( 5 ):401–403. doi: 10.1038/nmeth.3325 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]168.
Cano-Rodriguez D, Gjaltema RA, Jilderda LJ, ve diğerleri. H3K4Me3’ün yazılması epigenetik susturulmayı sürdürülebilir ancak bağlama bağlı bir şekilde aşar . Nat Commun . 2016; 7 :12284. doi: 10.1038/ncomms12284 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]169.
Lei Y, Zhang X, Su J, ve diğerleri. Tasarlanmış bir dCas9-MQ1 füzyon proteini kullanılarak canlı organizmada hedeflenen DNA metilasyonu . Nat Commun . 2017; 8 :16026. doi: 10.1038/ncomms16026 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]170.
Kwon DY, Zhao YT, Lamonica JM, Zhou Z. Sentetik bir CRISPR-Cas9 tabanlı HDAC kullanılarak lokus-spesifik histon deasetilasyonu . Nat Commun . 2017; 8 :15315. doi: 10.1038/ncomms15315 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]171.
Lorsch ZS, Hamilton PJ, Ramakrishnan A, ve diğerleri. Stres dayanıklılığı, prefrontal kortekste Zfp189 tarafından yönlendirilen bir transkripsiyonel ağ tarafından desteklenmektedir . Nat Neurosci . 2019; 22 ( 9 ):1413–1423. doi: 10.1038/s41593-019-0462-8 [ PMC ücretsiz makale ] [ PubMed ] [ CrossRef ] [ Google Akademik ]
/storage.evrimagaci.org%2Fold%2Fcontent_media%2Fd1a8032e99451f2f261e178218df8a3d.jpg)