Soyut
Biyolojik çeşitliliğin zaman içinde nasıl değiştiğini anlamak, evrimsel biyolojinin temel hedeflerinden biridir. Ancak, geçmiş biyolojik çeşitlilik tahminleri, örnekleme sapmalarını düzeltirken tahminleri ayarlamak için en son istatistiksel yöntemler uygulandığında bile, fosil kayıtlarının içsel eksikliği tarafından zorlanmaktadır. Burada, zaman içinde küresel veya bölgesel ölçeklerde zenginliği çıkarırken, mekansal, zamansal ve taksonomik örnekleme çeşitliliğini de içeren biyoçeşitliliğin stokastik simülasyonlarına ve derin öğrenme modeline dayalı bir yaklaşım geliştiriyoruz.
Yöntemimiz, özellikle büyük mekansal ölçeklerde, simüle edilmiş veri kümeleri arasında alternatif yaklaşımlardan daha iyi performans göstererek, çok çeşitli koruma senaryoları altında sağlam paleoçeşitlilik tahminleri sağlar. Yöntemimizi, farklı taksonomik ve zamansal kapsamdaki iki ampirik veri kümesine uyguluyoruz: deniz hayvanlarının Permiyen-Triyas kaydı ve hortumluların Senozoyik evrimi. Tahminlerimiz, deniz kayıtlarındaki iki kitlesel yok oluşun gözden geçirilmiş niceliksel değerlendirmesini sunuyor ve hortumluların Afrika’dan yayılmalarının ardından hızla çeşitlendiğini ve Pleistosen’de çeşitlilikte %70’ten fazla bir düşüş olduğunu ortaya koyuyor.
Devamını oku: DeepDive: Derin öğrenmeyi kullanarak zaman içinde küresel biyoçeşitlilik modellerini tahmin etmegiriiş
Zaman içinde biyolojik çeşitlilikteki değişimler, türlerin çeşitlenmesi ve yok oluşunun temel mekanizmalarını yansıtır; dinamiklerini tahmin etmek, yaşam tarihini anlamak için çok önemlidir. Fosil kayıtları, tahminlerin dayandırılacağı ampirik kanıtlar sunarak, yok olma, iyileşme, yayılma ve faunal ve floral dönüşüm süreçlerine ilişkin içgörüler sunarken, biyolojik çeşitlilik değişiminin itici güçlerinin yorumlandığı bağlamı belirler 1 , 2 , 3 , 4 , 5 . Evrimsel biyolojideki temel sorular, örneğin biyolojik çeşitliliğin küresel sınırlarının olup olmadığı 6 , 7 veya çevresel değişim, kitlesel yok oluşlar ve biyotik etkileşimler tarafından şekillendirilen biyolojik çeşitliliğin nasıl evrimleştiği 8 , 9 , 10 , 11 , 12 derin zamanda çeşitlilik kalıplarını çıkarsama yeteneğimize dayanır.
Fosil kayıtları geçmiş biyoçeşitlilik dinamiklerinin en doğrudan kanıtı olsa da, çok sayıda koruma ve örnekleme önyargısından etkilenir. Bu önyargılar, örnekleme ve sayısallaştırma çabalarındaki çeşitliliği, fosil alanlarının erişilebilirliğini, farklı organizmaların, habitatların ve coğrafi bölgelerin içsel koruma potansiyelini ve jeolojik tarihlerini yansıtır 13 , 14 , 15 , 16. Sonuç olarak, fosil kayıtları geçmiş biyoçeşitliliğin eksik bir örneğidir ve gerçek ve örneklenen çeşitlilik kalıpları arasında çıkarımsal bir uyumsuzluğa yol açan zamansal, mekansal ve taksonomik heterojenliklerle doludur 14 , 17 , 18 .Bu sorunu ele alma çabaları, örnekleme yoğunluğundaki değişimi hesaba katarak zaman içinde çeşitlilik yörüngelerini tahmin eden bir dizi yaygın olarak kullanılan yöntemle sonuçlanmıştır. Bunlar arasında birkaç seyreltme yöntemi 19 , 20 , 21 , 22 , Poisson örnekleme süreçlerine dayalı maksimum olabilirlik veya Bayes modelleri 23 , 24 ve alt sınır zenginlik ekstrapolatörleri 20 , 25 , 26 , 27 yer almaktadır. Bu yöntemler çoğunlukla zaman içinde koruma oranlarındaki değişimi hesaba katmaya odaklanırken, örneklemenin coğrafi kapsamındaki, zamansal süresindeki veya çevresel temsilindeki değişimi ele almazlar 28. Bu nedenle, fosil kayıtlarının mekansal ve zamansal heterojenliği, örnekleme standardizasyonundan sonra bile küresel biyoçeşitlilik tahminlerini her zaman engellemeye devam etmektedir 29 ve sığ deniz fosil kayıtlarının yakın zamanda yapılan bir analizi, mekansal örnekleme heterojenliğinin standartlaştırılmış zenginlik tahminlerindeki değişikliklerin %50-60’ını açıkladığını bulmuştur 14 . Fosil kayıtlarındaki bu önyargıların kapsamının giderek daha fazla anlaşılması, araştırma çabalarında hem mekan hem de zaman yoluyla çeşitliliğin daha mekansal olarak açık çalışmalarına doğru bir kaymayı gerektiriyor 17 , 30. Son çabalar bu kaymayı yansıtıyor 24 , 26 , 31 , 32 ve mekansal-zamansal önyargıların etkisine ilişkin anlayışın bunlarla mücadele yöntemleriyle veya makul biyoçeşitlilik modellerini simüle eden teorik modellerle uzlaştırılması yönünde ilerleme sağlıyor 33 , 34 , 35. Yine de, mekansal olarak açık yöntemler çoğunlukla bölgesel çeşitlilikteki zaman içindeki ve bölgeler arasındaki değişimi tahmin etmek için kullanılmış 14 , 24 , 29 , 36 ve küresel çeşitlilikteki değişimi tahmin etme sorununa doğrudan bir çözüm sunmamıştır. Taksonomik önyargılar, taksonların değişken korunması ve örneklemesinden kaynaklanan yaygın sorunlara rağmen mevcut yöntemlerin mevcut aralığında hesaba katılmamıştır 18 , 37 , 38 , 39 . Bazı modeller soylar arasında farklı koruma oranlarını hesaba katabilirken 24 , 40 , bu önyargıların gözlemlenmeyen soylar üzerindeki etkilerini açık bir şekilde hesaba katmazlar.
Burada, derin öğrenmeye dayalı çıkarımla mekanik bir simülasyon yaklaşımını birleştirerek fosil verilerinden zaman içinde biyoçeşitliliği tahmin etmek için bir çerçeve sunuyoruz. DeepDive (Derin Öğrenme Çeşitlilik Tahmini) adlı yaklaşımımızın performansını kapsamlı simülasyonlar aracılığıyla değerlendiriyoruz ve güçlü zamansal, mekansal ve taksonomik örnekleme önyargılarının varlığında bile çeşitlilik yörüngelerini doğru bir şekilde tahmin etme yeteneğini gösteriyoruz. Daha sonra DeepDive ile eğitilmiş modelleri kullanarak iki hayvan grubu için küresel biyoçeşitlilik dinamiklerini tahmin ediyoruz: Geç Permiyen’den Erken Jura’ya kadar olan deniz hayvanları 24 ve memeli kladı Proboscidea 41 .
Sonuçlar
Zaman içinde biyolojik çeşitliliği çıkarsamaya yönelik bir yaklaşım
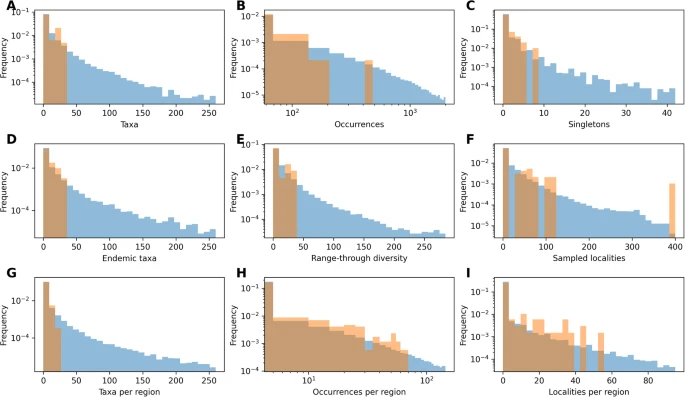
Biyoçeşitlilik yörüngelerini tahmin etmek için iki ana modülden oluşan bir çerçeve geliştirdik: 1) sentetik biyoçeşitlilik ve fosil veri kümeleri üreten bir simülasyon modülü ve 2) zaman içinde çeşitliliği tahmin etmek için fosil verilerini kullanan bir derin öğrenme çerçevesi (Şekil 1 ). Simülasyon modülü, türleşme, yok olma, fosilleşme ve örnekleme süreçlerine ilişkin anlayışımızı yansıtan veri kümeleri üretmek üzere tasarlanmıştır. Simülatör, geniş bir bölgesel heterojenlik yelpazesini kapsayan gerçekçi çeşitlilik yörüngeleri üretir (örn. Ek Şekil 1 ). Simüle edilen veriler ayrıca fosil oluşumlarını ve bunların ayrı coğrafi bölgelerdeki ve zaman içindeki dağılımlarını da içerir ve bunlar geniş bir yelpazedeki mekansal, zamansal ve taksonomik örnekleme önyargılarını yansıtacak şekilde üretilir (Ek Şekil 2 ). Bu veriler , ikinci modülde uygulanan tekrarlayan bir sinir ağına (RNN 42 , 43 , 44 , 45 , 46 ) dayalı bir derin öğrenme modelini eğitmek için kullanılır . RNN, küresel çeşitlilik yörüngesini tahmin etmek için fosil kayıtlarından çıkarılan tekillerin sayısı veya bölge başına yerellik sayısı gibi özellikleri kullanır. Simülasyon modülünde ayrıntılı olarak açıklanan biyocoğrafik bilgiler bu özelliklerde yansıtılır. Modelin birçok farklı veri kümesi üzerinde eğitilmesiyle RNN’nin parametreleri fosil kaydının genel özelliklerini öğrenir ve farklı evrimsel senaryolar ve örnekleme önyargıları boyunca biyoçeşitlilik yörüngelerini tahmin etmek üzere optimize edilir.
Şekil 1: DeepDive iş akışının gösterimi.
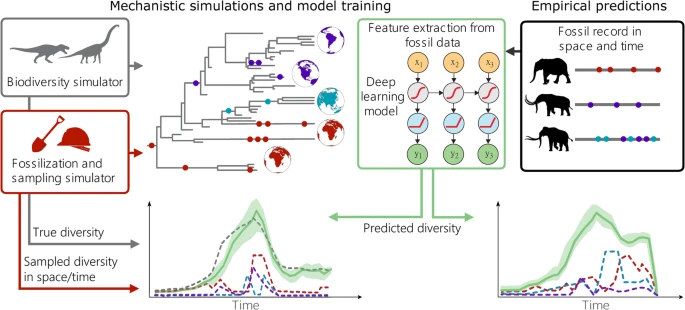
DeepDive yaklaşımı ayrıca, iki deneysel çalışmayla gösterdiğimiz gibi, zamansal ve biyocoğrafik kısıtlamalar ekleyerek eğitim simülasyonları kümesini belirli deneysel kladlara göre uyarlamamıza olanak tanır. Bu, eğitilmiş modellerimizi, jeolojik kayıtlardan ve modellerden bilinen kara kütleleri veya okyanus havzaları arasındaki bağlantıdaki değişiklikler veya daha önce çıkarılmış kitlesel yok oluş olayları gibi, ilgi duyulan kladın fosil kayıtlarında doğrudan gözlemlenemeyebilecek a priori deneysel bilgiye dayanarak bilgilendirebileceğimiz anlamına gelir. Örneğin, Proboscidea benzeri model için özel eğitim simülasyonları, kladın zaman diliminde kökeni ve kladın bu kıtalara taşındığı tahmini zamanlarda yalnızca simüle edilen türler tarafından işgal edilmesine izin verilen 5 kıta bölgesi ile başlama gereksinimini ve en az üç mevcut türle ilgili bir son kısıtlamayı içerir. Biyocoğrafyaları değiştirmek, simülasyon parametrelerini değiştirerek tanımlanabilir, örneğin iki bölge birleştiğinde veya taksonlar yeni bir bölgeye göç ettiğinde dağılım bağlantısını artırarak veya değişken dağılım ve bağlantı oranları kullanılabilir. Deniz verileri için özel eğitim simülasyonlarında, simülasyonlar birçok taksonla başlar ve biter ve çalışmanın zaman çerçevesinde bilinen iki büyük kitlesel yok oluş olayına göre kitlesel yok oluş olasılığına sahiptir. Simüle edilmiş veri kümelerindeki parametrelerin dağılımı, ampirik oluşum verilerindekilerle karşılaştırılabilir ve ampirik verilere dayalı olarak beklenen parametre aralığının modelin öğrenme fırsatı bulduğu aralıkta olduğundan emin olunabilir. Bu şekilde, simülasyonları özelleştirmek ve dolayısıyla bu veriler üzerinde eğitilen modelleri, eğitim kümelerinin ampirik durumla ilişkilendirilebilir olduğundan emin olarak çok çeşitli olası evrimsel senaryolara göre özelleştirmek mümkündür. Ek Tablo 1’de özetlenen Parametreli ve notasyonlu Yöntemler bölümünde daha fazla ayrıntı verilmiştir .
DeepDive’ın Performansı
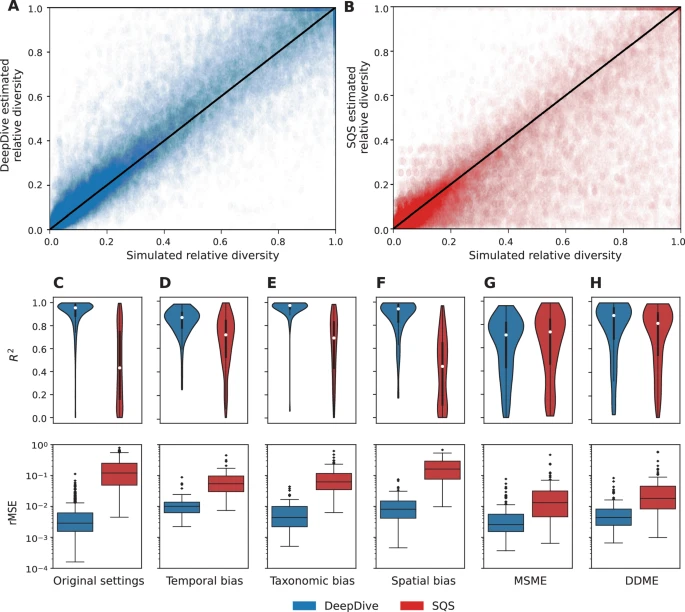
DeepDive tarafından eğitilen modellerin performansını, çok çeşitli çeşitlendirme senaryolarını kapsayan kapsamlı simülasyonlar aracılığıyla doğruladık, modelleri optimize etmek için eğitim setleri oluşturduk ve performanslarını bağımsız olarak oluşturulan doğrulama ve test setleri üzerinde değerlendirdik. Hem simüle edilen hem de tahmin edilen çeşitlilik yörüngelerini 0 ile 1 arasında bir aralığa yeniden ölçeklendirdikten sonra hesaplanan yeniden ölçeklenmiş ortalama karesel hatayı (rMSE) ve modelin performans ölçütleri olarak dönüştürülmemiş çeşitliliğe ( R 2 ) uygulanan belirleme katsayısını kullandık. Bunlar, tahmin edilen biyoçeşitliliğin simüle edilen yörüngeyle ne kadar doğru bir şekilde eşleştiğini göreceli bir ölçekte değerlendirmemizi sağladı. Simülasyonlarımızda göreceli çeşitliliğe odaklansak da, bu, Hissedar Çoğunluğu Alt Örneklemesi (SQS) alt örnekleme yaklaşımıyla daha adil bir karşılaştırmaya izin verdiği için, DeepDive’ın çıktısı zaman içinde mutlak çeşitliliği nicelleştirir. Bu nedenle, DeepDive’ın doğruluğunu nicelleştirmek için simüle edilen ve tahmin edilen çeşitlilik yörüngeleri arasındaki ortalama karesel hatayı (MSE) da rapor ediyoruz.
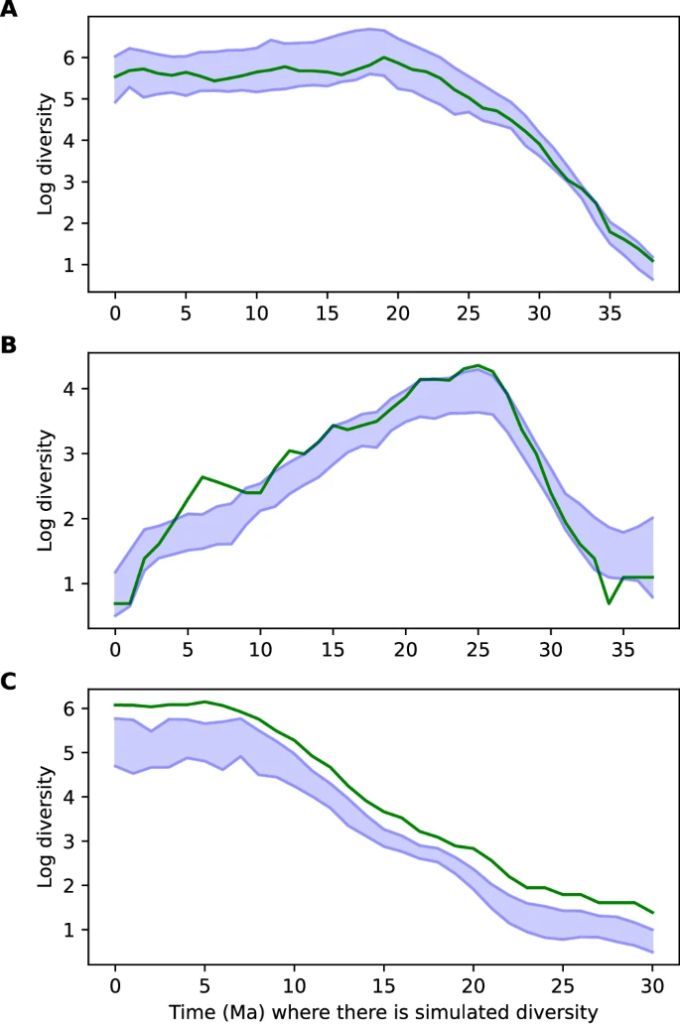
Optimum yapılandırmayı değerlendirmek için bir dizi model mimarisini test ettik (Ek Tablo 2 ) ve çoğu modelin 0,114-0,132 civarındaki doğrulama MSE aralığında ve 0,197-0,229 arasındaki test MSE aralığında benzer şekilde iyi performans gösterdiğini bulduk. Bu, tahminlerin RNN çerçevemiz içindeki bir dizi parametrelendirme boyunca tutarlı olduğunu gösterir. Tahminler, çoğu durumda simüle edilenlerle yakından eşleşerek, çok çeşitli yörünge senaryolarında doğruydu (Ek Şekil 3 ve Şekil 2 A). Tahminlerle ilişkili belirsizliği ölçmek için bir Monte Carlo bırakma katmanı 46 ekledik ve her model için birden fazla tahmin yaptık. Çeşitlilik tahminleri etrafında %95 güven aralıkları elde etmek için farklı eğitilmiş modeller arasında Monte Carlo bırakma tahminlerini birleştirdik. Tahminlerin simüle edilmiş biyoçeşitlilik değerlerinin kalıplarına oldukça benzediğini, simüle edilmiş değerlerin test seti simülasyonları boyunca ihmal edilemez bir zaman diliminde Monte Carlo bırakma yoluyla elde edilen %95 güven aralıklarında olmadığını bulduk. Özellikle, medyan kapsam, yani simüle edilmiş değerlerin %95 aralığına dahil edilen oranı, test simülasyonları boyunca %66 idi (Şekil 3 ), bu da Monte Carlo bırakmada gerçek belirsizlik aralıklarını küçümseme eğilimini yansıtmaktadır 47 .
Şekil 2: DeepDive yaklaşımı (mavi) , farklı önyargı koşulları altında hataları azaltabilir ve
SQS’ye (kırmızı) göre R2’yi iyileştirebilir.
Şekil 3: Simüle edilmiş çeşitlilik eğrilerinde DeepDive tahminlerinin kapsamı.
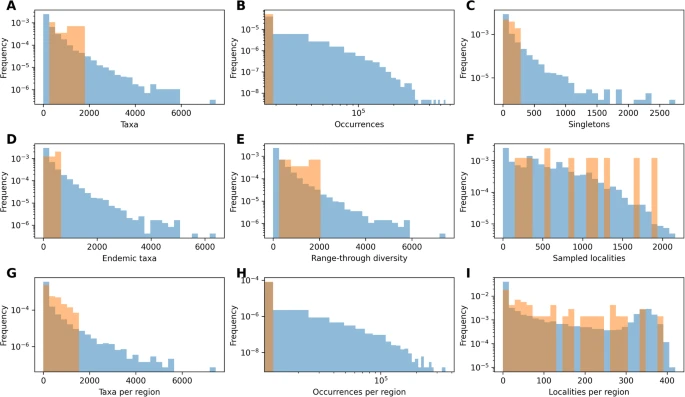
DeepDive ile eğitilen modellerin performansının, fosil kayıtlarının kalitesindeki değişimden nasıl etkilendiğini değerlendirdik. Bu değişimleri (1) bütünlük (en az bir fosil varlığına sahip türlerin oranı); (2) korunma oranı (zaman dilimi başına soy başına ortalama fosil kaydı sayısı); (3) örneklenen tür sayısı; (4) taksonların süresi; (5) kladların süresi olarak belirledik. Beklendiği gibi DeepDive tahminleri, yüksek korunma oranlarına ve daha fazla örneklenen türe sahip daha eksiksiz veri setlerinde en doğruydu. Düşük hata (rMSE < 0,01), bütünlüğün 0,2’yi aştığı (yani fosil kayıtlarında türlerin %80’ine kadarının örneklenmediği) çoğu simülasyonda bulundu ve daha düşük bütünlük seviyelerinde daha yüksek hataya sahip veri setlerinin sıklığı arttı (Ek Şekil
4 A). Benzer bir eğilimi, daha yüksek korumaya sahip veri kümelerinde rMSE’deki değişimin en düşük olduğu (Ek Şekil
4 B) ve hata büyüklüğünün yalnızca ~200’den az örneklenen türe sahip veri kümelerinde 0,01’in önemli ölçüde üzerinde arttığı (Ek Şekil
4 C) koruma oranlarında da gözlemledik. Bir kladın analizin zaman çerçevesinin sonunda neslinin tükenmesi veya varlığını sürdürmesi doğruluk üzerinde
önemli bir etkiye sahip değildir (Ek Şekil 4 D). Tahminler, ortalama olarak kısa ömürlü türlerle karakterize edilen veri kümelerinde daha fazla hataya açık olma eğilimindeyken, klad süresi ile MSE puanları arasında net bir ilişki bulamadık (Ek Şekil 4 E–F).
SQS ile karşılaştırma
DeepDive’ı fosil verilerinden çeşitlilik yörüngelerini tahmin etmek için en yaygın olarak uygulanan yöntemlerden biri olan SQS 19 , 27 ile karşılaştırdık 48 , 49 , 50 , 51. SQS göreceli çeşitliliği tahmin ettiğinden, performanslarını karşılaştırmak için rMSE ve R2 ölçümlerini kullandık. Analizde, DeepDive ile eğitilen modeller, test simülasyonları boyunca birden fazla büyüklük sırasına göre daha düşük göreceli medyan rMSE ve daha yüksek bir medyan belirleme katsayısı (0,958’e karşı 0,432) ile SQS’den daha iyi performans gösterdi (Şekil 2 ). Özellikle, DeepDive tahmini verilerdeki boşluklara karşı daha dayanıklı görünüyor ve hem düzgün hem de ani çeşitlilik değişiklikleri dahil olmak üzere genel biyoçeşitlilik dinamiklerini daha iyi yakalıyor gibi görünüyor (Ek Şekil 3 ). Hem DeepDive hem de SQS tahminleri düşük düzeydeki tamamlanma ve koruma oranlarından olumsuz etkilenirken (Ek Şekil 5 ), DeepDive örnekleme senaryolarının tüm spektrumunda önemli ölçüde daha doğru tahminler sürdürdü (Ek Şekil 4 ).
Her iki yöntemi de kullanarak, güçlü zamansal, taksonomik ve mekansal önyargı koşullarını ve başlangıç eğitim setine açıkça dahil edilmemiş çeşitlilik değişim modellerini yansıtan beş ek simüle edilmiş veri setini (her durumda 100 simülasyon) test ettik: kitlesel yok oluşun izlediği çeşitlilik bağımlılığı ve çoklu kitlesel türleşme ve kitlesel yok oluş olaylarının olduğu simülasyonlar (Şekil 2 D–H). Belirleme katsayıları, örnekleme oranlarının zamansal değişiminin her iki yöntem altında da etkili bir şekilde düzeltilebileceğini gösterdi, ancak DeepDive tahminleri daha doğruydu. Taksonomik ve daha büyük ölçüde mekansal önyargılar, iki yöntem arasında önemli ölçüde daha yüksek bir tutarsızlığa yol açtı. Güçlü mekansal önyargılar durumunda, medyan R2 , DeepDive için 0,9’u aştı ve SQS tahminleri için yaklaşık 0,25’e düştü. Eğitim setinde nadir bulunan desenler analiz edildiğinde, modelin performansı R2 açısından SQS’ye benzer seviyelere düşerken , yine de önemli ölçüde daha düşük rMSE’yi korur (Şekil 2 G–H). Ancak, yeniden eğitilen bir modelin eğitim setine kitle türleşmesi ve çeşitlilik bağımlılığının dahil edilmesi doğruluğu artırır (Ek Şekiller 6 – 7 ). Çeşitlilik bağımlılığı ve ardından kitlesel yok oluş gibi desenlerin dahil edilmesi, kitlesel yok oluş olaylarının tahmini etrafındaki hataları da azaltabilir (Ek Şekil 8 ).
Çeşitliliğin deneysel verilerden tahmin edilmesi
DeepDive yaklaşımını farklı taksonomik ve coğrafi kapsamdaki iki ampirik veri kümesine uyguluyoruz ve tahminlerimizi zaman içinde göreceli ve mutlak biyoçeşitlilik değişimlerini değerlendirmek için kullanıyoruz. Birincisi, Geç Permiyen’den Erken Jura’ya kadar uzanan deniz hayvanlarının cins düzeyindeki veri kümesidir ve 5312 cins briyozoa, knidarya, brakiopod, yumuşakça, ekinoderm, foraminifer, eklembacaklı, kordalı, porifer ve retaryan’da 71.386 oluşumu içerir (bkz. “Yöntemler”). İkincisi, modern filler ve soyu tükenmiş akrabalarını içeren Proboscidea takımının tür düzeyindeki fosil veri kümesidir ve 180 türde 2104 oluşumu içerir (bkz. “Yöntemler”).
Her veri setindeki deneysel zaman bölmelerinin sayısını ve süresini yansıtan ve mümkün olduğunda çeşitlilik ve mekansal kısıtlamaları içeren simüle edilmiş veri setlerine dayalı modeller eğittik (daha fazla ayrıntı için “Yöntemler”e bakın). Simüle edilmiş veri setlerindeki özellikler, deneysel veri setlerinde gözlemlenen özelliklerin aralığını kapsar (Şekiller 4 – 5 ). Modellerin doğruluğunu değerlendirdikten sonra (Ek Tablolar 3 – 4 ), iki veri seti için çeşitlilik yörüngelerini tahmin ettik.
Şekil 4: DeepDive simülasyonlarının denizsel deneysel verilere göre değişimi.
Şekil 5: DeepDive simülasyonlarındaki hortumlu hayvanlara ait deneysel verilere göre değişim.
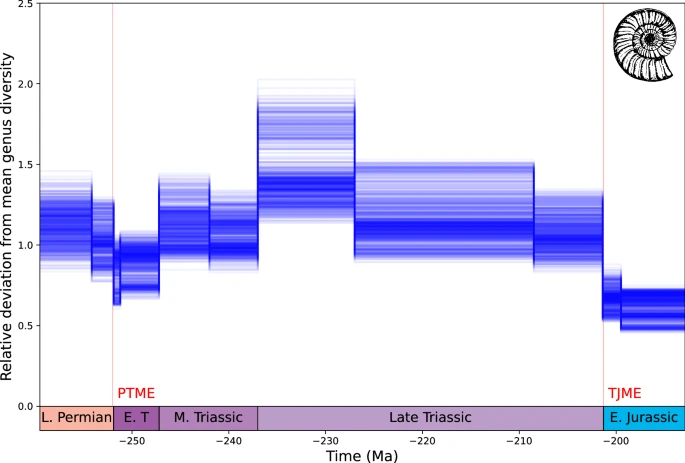
Deniz çeşitliliği Geç Permiyen ile en erken Triyas arasında azalır ve Permiyen-Triyas kitlesel yok oluşu (PTME) sırasında cinslerin %58’ine kadar kayıp yaşanır (Şekil 6 ), ancak daha düşük büyüklükler de mümkündür (ortalama %24 cins kaybı). Erken Triyas’ta cins sayısı kurtarıldı ve Orta Triyas’ta PTME öncesi seviyeleri aştı. Çeşitlilik Geç Triyas boyunca daha kademeli olarak azalır ve ardından cinslerin %66’sına kadar kayıpla (ortalama %42 cins kaybı) Triyas-Jura sınırında daha keskin bir düşüş görülür.
Şekil 6: DeepDive’ın Permo-Triyas denizel cinsleri için tahmin ettiği çeşitlilik.
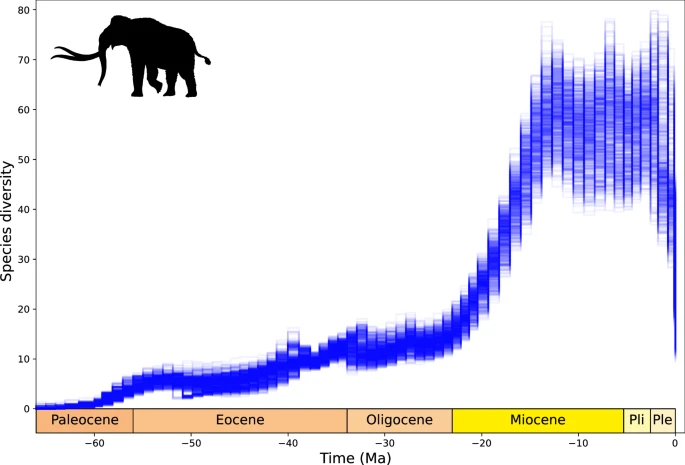
Hortumlular kladının çeşitlenme geçmişini Erken Senozoyik’teki (66-60 Ma) kökeninden bu yana tahmin ettik ve zaman içinde kademeli bir çeşitlilik artışı olduğunu ve Miyosen’in başlangıcında tahmini tür zenginliğinin 10-20 türe ulaştığını çıkardık (Şekil 7 ). Erken Miyosen’de (23-15 Ma) çeşitlilikte dik bir artış tespit ettik ve bu da Orta ila Geç Miyosen’de Dünya’da dolaşan 35-78 kadar çağdaş fil türüne yol açtı. Çeşitlilik, çalışma aralığının sonunda 10-27 türle Pleyistosen boyunca çökmeden önce Pliyosen’de ve Pleyistosen’in başında nispeten yüksek ve değişken kaldı (ortalama %65 tür kaybı, maksimum %87, minimum %32). Bu yeniden yapılanmanın ışığında, üç türün günümüzdeki çeşitliliğinin, Pleyistosen’den (2,58 milyon yıl önce) bu yana %70’in üzerinde tür kaybından kaynaklandığı görülmektedir (maksimum kayıp %89, ortalama %85).
Şekil 7: DeepDive’ın Proboscidea için tahmin ettiği çeşitlilik.
Kaynak ve devamına Buradan ulaşabilirsiniz.