Soyut
Trilyonlarca mikroorganizma, topluca mikrobiyom olarak bilinir, vücudumuzda yaşar ve bağırsak mikrobiyomu biyomedikal araştırmalarda özel ilgi odağıdır. Baskın virom bileşenleri olan bakteriyofajlar, konakçıları standart bakteri koduyla enfekte ederken alternatif genetik kodlara geçmek için baskılayıcı tRNA’ları kullanabilir (örneğin, UAG durdurma kodonu glutamine yeniden atanır).
Ancak, bu geçişi neyin tetiklediği ve bakteriyofajın konakçısını nasıl manipüle ettiği yeterince anlaşılmamıştır. Burada, yeniden kodlanmış lizis ve yapısal faj genlerinin ifadesine yol açan kod geçişinde potansiyel olarak yer alan minimal hepatit delta virüsü (HDV) benzeri ribozimlerin bir alt grubunun – teta ribozimlerinin – keşfini bildiriyoruz. HDV benzeri kendi kendine ayrılma davranışlarını in vitro gösteriyoruz ve bunları bildirilmemiş bir bağlamda, genellikle ayrılma yerleri tRNA’lara bitişik olarak konumlanmış olarak buluyoruz; bu da viral tRNA olgunlaşmasında ve/veya düzenlenmesinde bir rol olduğunu gösteriyor. İlişkili her beş tRNA’dan biri baskılayıcı bir tRNA’dır ve bu da hipotezimizi daha da güçlendirir. tRNA ile ilişkili teta ribozimlerinin muazzam bolluğu – 1753 benzersiz örnek sunuyoruz – genellikle tRNA 3′ uçlarını işleyen büyük enzimlere bir alternatif olarak küçük ribozimlerin önemini vurgular. Keşfimiz, küçük HDV benzeri ribozimlerin kısa biyolojik işlevleri listesini genişletiyor ve muhtemelen belirli yeniden kodlanmış bağırsak bakteriyofajlarının kod değişiminde yer alan daha önce bilinmeyen bir oyuncuyu tanıtıyor.
giriiş
Ribozimler her yerde bulunur ve peptidil transferaz aktivitesi 1 ve tRNA olgunlaşması için gerekli transesterifikasyon adımları 2 ve ökaryotik mRNA eklenmesi 3 dahil olmak üzere yaşamın tüm alanlarındaki temel biyolojik süreçlere katılırlar . Küçük ribozimler (<200 nükleotid; nt) kendi kendine kesme ve/veya -ligasyonla sınırlıdır ancak dizi, yapı ve biyolojik işlevlerde dikkate değer ölçüde çeşitlidirler 4 , 5 , 6 , 7 , 8 . İyi çalışılmış bir örnek, oldukça korunmuş, yuvalanmış çift psödoknotlu bir yapıya sahip ancak birincil dizide önemli değişkenliğe sahip olan HDV benzeri ribozimler ailesidir (delta benzeri ribozimler, drzs) 9 , 10 , 11 (Şekil 1a ). Belirli drz örneklerinin biyolojik işlevleri bilinmesine rağmen 12 , 13 , 14 , 15 , 16 , 17 çoğunluğu, özellikle minimal varyantlar, daha az anlaşılmıştır. İlk olarak metagenomik örneklerde tanımlanan minimal drz’lerde P4 alanı yoktur 18 (Şekil 1a ) ve kökenleri (ökaryotik, bakteriyel veya viral) ve biyolojik işlevleri henüz belirlenmemiştir.
Şekil 1: HDV benzeri ribozimler (drz’ler) ve bakteriyofajlarla ilişkileri.
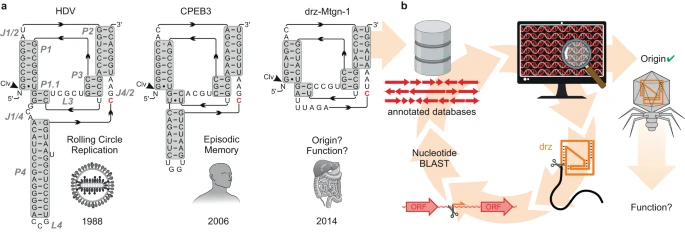
Burada, memeli bağırsağındaki Caudoviricetes bakteriyofaj genomlarında sıklıkla viral tRNA’larla ilişkilendirilen minimal drz’lerin keşfini ve in vitro doğrulamasını bildiriyoruz ve bunları teta ribozimleri (Θrz’ler) olarak adlandırıyoruz. Bağırsak viromu esas olarak bakteriyofajlardan (>%90) oluşur ve giderek artan bir şekilde insan sağlığı ve hastalıklarıyla ilişkilendirilmektedir 19 , 20 , 21 , 22 , bu da son yıllarda çok sayıda dizi veri tabanının eklenmesine yol açmıştır. Bu veri tabanlarının dizi analizleri, bazı bakteriyofajların yeniden kodlandığını ortaya koymuştur: belirli bir durdurma kodonunun standart bir amino aside yeniden atandığı genetik kodlar kullanırlar 23 . Kehribar durdurma kodonunun (UAG) glutamine (kod 15) yeniden kodlandığı bir örnek, yakın zamanda deneysel olarak doğrulandı 24 . Viral baskılayıcı tRNA’lar (tRNA Sup ), alternatif genetik kodların çevirisinde merkezi oyunculardır ve yeniden kodlanmış fajlar tarafından konak lizini başlatmak için kullanılabilecek bir araçtır. Ancak, bakteriyofajlardaki litik-lizojenik geçişin kesin mekanizması henüz tam olarak karakterize edilmemiştir.
Bu çalışmada, tRNA ile ilişkili Θrz’lerin viral tRNA olgunlaşmasında rol oynadığını ve yeniden kodlanmış bakteriyofajların bir alt kümesinde yeniden kodlanmış durdurma kodonları içeren geç faz lizinin ve yapısal genlerin ifadesini destekleyebileceğini, hatta potansiyel olarak faj lizini tetikleyebileceğini ileri sürüyoruz. Bulgularımız, drz’lerin ilgi çekici dünyasına ve biyolojik önemlerine dair içgörüler sunmaktadır.
Sonuçlar
Metagenomik minimal drz’lerin viral kökeni
Minimal drz “drz-Mtgn-1” metagenomik bir örnekte tanımlandı 18 , ancak kökeni ve biyolojik işlevi belirsizliğini koruyor (Şekil 1a ). Bu bilgi boşluğu ve drz’nin iki değerlikli metal iyonlarıyla olan benzersiz davranışı bizi meraklandırdı ve halka açık dizi veritabanlarında nükleotid dizisine dayalı bir arama yürüttük 25 . Bu arama, diğerlerinin yanı sıra, drz-Mtgn-1’in birkaç çift sarmallı DNA (dsDNA) bakteriyofajına atanmasına yol açtı ( Caudoviricetes , Şekil 1b ).
Drz sekonder yapısının birincil diziden ziyade korunması nedeniyle, yaklaşımımızı yayınlanmış bir drz dizisiyle temel bir yerel hizalama arama aracı (BLAST 26 ) kullanmaktan, 18 numaralı referansla minimal motifi kullanan RNArobo 27 ile motif tabanlı bir aramaya çevirdik. İlk aramalar, bakteriyofaj genom veritabanlarında 28 , 29 60’ın üzerinde minimal drz dizisiyle sonuçlandı ve üç sonuç ortaya çıktı: (i) minimal drz dizileri başlangıçta yalnızca insan bağırsağı metagenomik verilerinden derlenen bakteriyofaj genomlarında tespit edildi, (ii) isabetler yakındaki açık okuma çerçeveleriyle (ORF; çoğunlukla bilinmeyen işlevlere sahip varsayımsal proteinler) ilişki gösterdi ve potansiyel olarak biyolojik işlevleri hakkında fikir verdi (Ek Şekil 1 ) ve (iii) minimal drz’ler daha önce düşünülenden daha yaygındır; 2014’te gerçekleştirilen tam ölçekli bir aramada keşfedilen birkaç isabete kıyasla iki veritabanında yapılan ilk aramada keşfedilen düzinelerce isabet vardır 18 .
tRNA ile ilişkili teta ribozimlerinin (Θrzs) keşfi
İlk kategorizasyonumuzdaki isabetlerin bir alt kümesi, özellikle faj tRNA genlerine bitişik minimal drz’ler dikkatimizi çekti. tRNA’nın 3′ ucundaki ribozim kesme bölgesinin (G1; Şekil 1a ) konumu, tRNA olgunlaşmasında daha önce bilinmeyen bir biyolojik işlevi olduğunu düşündürmektedir. Bu nedenle bu örneklere odaklandık ve arama motifimizi buna göre geliştirdik. Çeşitli ortamlardan yakın zamanda ek açıklama eklenmiş sekiz viral veritabanı 23 , 29 , 30 , 31 , 32 , 33 , 34 , 35 seçtik (Ek Tablo 1 ) ve aynı veritabanlarındaki tRNA motif aramalarıyla tüm sonraki isabetleri çapraz referansladık. Motif özgüllüğünü artırmak için, drz’lerin aktif bölgedeki 36 sitozin-urasil mutasyonu (CΔU) ile inaktive edildiğine dair önceki bulguları göz önünde bulundurarak, dahili kontroller olarak yanlış pozitif motifleri dahil ettik; bu pozisyonda 37 hiçbir kurtarma mutasyonu gözlenmedi . Her arama, dört farklı tanımlayıcı dosya ile gerçekleştirildi; katalitik sitozin kalıntısı sağlam olan bir aktif ribozim motifi (J4/2 bağlantısındaki ilk pozisyon) ve bu kalıntıda ikameler içeren üç yanlış pozitif motif (sırasıyla CΔA, CΔG ve CΔU; Şekil 2a ). İlk arama, aktif motifle toplam isabetlerin %50’sinden azını verdi ve bu da yüksek bir yanlış pozitif oranına işaret etti (Şekil 2b (i) ). Motifi, L4 halkasını ve J1/2 bağlantısını kısaltarak (Şekil 2b (i), (ii) ) ve ön fikir birliği dizilerine dayalı nükleotid kimliği kısıtlamaları uygulayarak yinelemeli olarak geliştirdik (>%97 koruma; Şekil 2b (iii) ). Son olarak, HDV benzeri yapısal motif 10 ile uyumlu olarak J4/2 bağlantısının son pozisyonuna bir serbestlik derecesi daha ekledik . İkincisi, yanlış pozitif oranını neredeyse sıfıra düşürürken, tespit edilen tRNA ile ilişkili ribozimlerin sayısını artırdı (Şekil 2b (iv) ). Bu rafine motiften elde edilen dizilere , tRNA’larla sık ilişkileri nedeniyle teta ribozimleri (Θrz) adı verilir .
Şekil 2: Açıklamalı bakteriyofaj veri tabanları kullanılarak teta ribozim (Θrz) motifinin optimizasyonu.
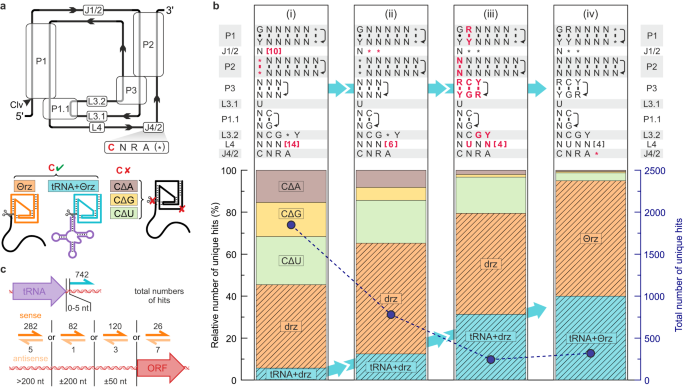
Optimize edilmiş motif kullanılarak, yukarıda belirtilen veritabanlarında 302 benzersiz Θrz dizisi tanımladık ve bunlardan 126’sı, ribozimin kesme noktasının bir tRNA’nın 3′-ucundan ±5 nt uzaklıkta olduğu tRNA ile ilişkili Θrz’ler olarak sınıflandırıldı. Analizimiz, 185 benzersiz tRNA/Θrz kombinasyonuyla sonuçlanan 152 ayrı Θrz-bitişik tRNA dizisi ortaya çıkardı. Birçok Θrz, birden fazla viral genomda mevcut olup toplam 1281 oluşuma sahiptir ve 742’si (%58) tRNA ile ilişkili Θrz olarak sınıflandırılmıştır (Şekil 2c , üst). Bu tRNA ile ilişkili Θrz’lerin 568’i (%77) ilgili tRNA’ya doğrudan bitişiktir (±1 nt). Geri kalan kısım (%23) için, tRNA uçlarının hesaplamalı tahminindeki küçük yanlışlıkların, tRNA ile Θrz kesme bölgesi arasındaki birkaç ek nükleotidi açıkladığını varsayıyoruz.
Θrz’lerin %80’inden fazlası insan veya hayvan bağırsağı bakteriyofajlarında bulundu, diğer ortamlardan izole edilen fajlarda hiç veya sadece birkaç isabet vardı (Ek Tablo 1 ). Bağırsakla ilişkili faj genomlarındaki zenginleşmenin tarafsız olduğunu doğrulamak için analizimize bakteri 38 ve ökaryotik genomlar 39’u (insan, fare ve protistler) dahil ettik. Bu veritabanları iki katından daha büyük olmasına rağmen, arama bakteri genomlarında yalnızca 12 Θrz isabeti ortaya koydu ve bunların hiçbiri bir tRNA ile ilişkili değildi. İlk manuel aramalarımızla tutarlı olarak, Θrz ve minimal drz dizilerinin yaklaşık %99’u insan bağırsağı viromu 40’taki baskın dsDNA virüsleri olan Caudoviricetes sınıfına ait bakteriyofajlarla sınırlıydı (Ek Şekil 2 ).
Mevcut olduğunda, silico konak tahminleri veritabanlarından çıkarıldı ve iki şubeyle sonuçlandı: Bakteriyel konakların baskın kısmı (%90 civarı) Bacteroidota şubesine aitken, geri kalanı Bacillota şubesinin bir parçasıdır . Bu şubelerin bazı üyeleri tRNA’larında yalnızca ara sıra bir CCA kuyruğu kodladığından 41 , bu temel özelliğin varlığı açısından faj tRNA’larını analiz ettik. İlginç bir şekilde, Θrz’lerle ilişkili tüm tRNA’ların yalnızca %4,5’i bir CCA kuyruğu kodlar. Ancak, Θrz içeren faj genomlarının %20,6’sının bir tRNA adenililtransferaz kodladığını ve bunun tRNA’lara transkripsiyon sonrası bir CCA kuyruğu eklenmesini sağladığını bulduk. Buna karşılık, analiz edilen tüm fajların yalnızca %0,08’i bu enzimi taşır, yani bu gen Θrz içeren fajlarda 250 kat artar. Θrz içeren faj genomlarının %54,5’inde tespit edilen benzer şekilde zenginleştirilmiş bir ORF (140 kat) “RNA ligaz, DRB0094 ailesi” olarak açıklanmıştır. Bu enzim, aminoasil-tRNA sentetazlarına benzeyen bir N-terminal modülüne bağlı bir C-terminal adenililtransferaz alanı içerir 42 . Bu nedenle, benzer bir işlevi yerine getirebileceğini ve ek olarak uygun amino asidi karşılık gelen tRNA’sına bağlayabileceğini öneriyoruz. Fajların geri kalan azınlığının tRNA olgunlaşmasının bu yönleri için muhtemelen konak tarafından kodlanan enzimlere güvendiğini varsayıyoruz.
tRNA ile ilişkili olmayan Θrz isabetleri ( n = 526) en yakın ORF’lerine göre kategorize edildi (Şekil 2c , alt). Çoğunluk ( n = 493) kodlamayan bölgelerde yer alıyor, çoğu en yakın açıklamalı ORF’den 200 nt’den daha uzakta yer alıyor. Bu Θrz isabetlerinin %96’sından fazlası en yakın yukarı veya aşağı akış geniyle aynı yönelimi (duyu) paylaşıyordu. Sadece 33 örnek kısmen veya tamamen bir ORF içinde yer alıyordu (intragenik). Ancak bu genomlardaki önerilen kodlama bölgeleri varsayımsal, doğrulanmamış ORF’lerdir ve bu nedenle yanlış pozitifler içerebilirler. Tanımlanan ribozimlerin HDV benzeri bir şekilde beklenen kendi kendine kesilmesini göz önünde bulundurarak, bunların kodlama bölgelerinin dışında yer almasını bekliyoruz. Bu nedenle, fajların çok yoğun kodlandığı bilindiği için, intragenik Θrz’lerin sıklığı yanlış pozitif oranımızı daha kesin bir şekilde (~%2,3) tahmin etmek için kullanılabilir, ancak yine de öngörülen ORF’ler içinde bu kadar düşük sayıda Θrz buluyoruz. Bir tRNA ile ilişkili olmayan kodlamayan bölgelerdeki Θrz’ler muhtemelen bilinmeyen biyolojik işlevlere hizmet eder ve gelecekteki çalışmalara konu olabilir. Ancak, tRNA ile ilişkili Θrz’ler hit havuzumuzun çoğunluğunu oluşturur ve biyolojik bir işlevin en açık göstergesini gösterir, bu da bizi in vitro doğrulama için bu alt gruba odaklanmaya yönlendirir.
tRNA ile ilişkili Θrz’ler in vitro aktiftir
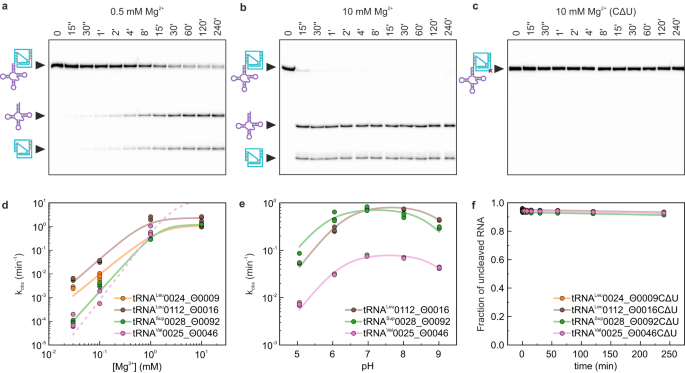
Drz’lerin iç transesterifikasyon mekanizması, bozulmuş bir p Ka ve koordineli bir Mg 2+ iyonu ile J4/2 bağlayıcısındaki temel bir sitozine dayanır ve bu da fosfat omurgasını bir hat içi saldırı için konumlandırır. Bu asit-baz katalizli reaksiyonun nötr pH 43 yakınında en verimli olduğu gösterilmiştir . tRNA ile ilişkili Θrz’lerin HDV benzeri kendi kendine ayrılmasını doğrulamak için, in vitro dört tRNA/Θrz çiftini seçtik ve inceledik (Ek Şekil 3 ve Ek Veri Dosyası 1 ). İlk motif aramamızda yüksek yaygınlıklarına dayanarak üç çift seçtik ve uzamış J4/2 bağlantısı nedeniyle tRNA Val 0025_Θ0046 çiftini (isimlendirme için Yöntemlere bakın) seçtik; bu, son motifte ek olarak tanıtılan serbestlik derecesiyle elde edilen isabetlerin gerçek pozitifler olarak deneysel olarak doğrulanmasını sağladı (Şekil 2b (iv) ).
Seçilen tüm örnekler in vitro Mg 2+ -bağımlı kendi kendine ayrılma aktivitesi gösterdi (Şekil 3a,b ve Ek Şekil 4a ). Görünür kendi kendine ayrılma hızı sabiti ( kobs ) , artan Mg 2+ konsantrasyonuyla tipik bir HDV benzeri sigmoidal davranış gösterdi . Yapılar arasında, tRNA/Θrz çifti tRNA Val 0025_Θ0046 , pH 7,0’da en yüksek kobs’yi gösterdi (>10 dk -1 10 mM Mg 2+ ‘da ; Şekil 3b,d ), diğer yapılar ise aynı Mg 2+ konsantrasyonunda (10 mM; Şekil 3d ve Ek Tablo 2 ) önemli ölçüde daha yavaş kobs gösterdi .
Şekil 3: Seçilmiş tRNA/Θrz çiftlerinin in vitro doğrulanması.
Kaynak ve devamına Buradan ulaşabilirsiniz.