SOYUT
Hastalıkla ilişkili genlerdeki amino asit değişimlerinin yapısal ve işlevsel etkisini etkileyen temel faktörlerden biri, arka plandaki genetik varyasyondur. Aynı gendeki birlikte evrimleşen pozisyonlar arasındaki epistatik etkileşimler, her bir bölgenin etkileşimde bulunduğu ortaklarla karşılıklı bağımlılığına yol açar. Bu etkileşimlerin rolünü destekleyen kanıtlar, insan dışı türlerin genomunda insan hastalıklarıyla ilişkili alellerin ortaya çıkmasıyla örneklendirilmiştir. Bu olguya çeşitli açıklamalar katkıda bulunabilse de, bir olasılık, patojenik etkiyi tamamen veya kısmen geri kazandırabilen ve protein yapısının ve düzgün işlevinin korunmasına katkıda bulunan telafi edici etkileşimli bölgelerin ortaya çıkmasıdır. Bu etkileşimler, burada belirli proteinler için literatür verilerinin analizi yoluyla yeniden ele alınan genetik bağlamlar yaratır; özellikle de doğuştan metabolizma hatalarında rol oynayan proteinlere odaklanır.
1 Giriş
Genotip ve fenotip arasındaki karmaşık korelasyonlar, özellikle tüm genom ve tüm ekzom dizilemesiyle keşfedilen çok sayıda varyant nedeniyle, tıbbi genetik alanı için en önemli zorluklardan bazılarını temsil eder. Tek bir varyantın sağladığı bilgi, genellikle ilgili protein üzerindeki etkisini tam olarak açıklamak için yeterli kabul edilebilir. Bununla birlikte, her varyantın, bireyin genetik arka planını birlikte oluşturan diğer varyantlarla birlikte bulunduğunun farkında olmak önemlidir. Hem Mendelyen hem de karmaşık hastalıklarda gözlemlenen genetik karmaşıklık, bir varyantın etkisinin diğer faktörlerden etkilenebileceğini ve bağlama bağlı olduğunu göstermektedir [ 1 – 3 ]. Bu bağlamda, bir genin tüm varyantlarını analiz etmek, özellikle fenotip-genotip korelasyonunun biyolojik karmaşıklığın diğer boyutlarını içerdiği durumlarda kapsamlı bilgi sağlayabilir [ 3 – 7 ].
Farklı varyantlar arasındaki etkileşime epistasis denir [ 8 ]. Terim şu anda yaygın olarak kullanılsa da [ 9 , 10 ], burada türler arası karşılaştırmalarla çıkarılan molekül içi etkileşimlerin özel bağlamına odaklandık. Üç boyutlu düzeyde varyantlar arasındaki etkileşim, bir proteinin düzgün çalışması için önemlidir [ 11-14 ] . Bu nedenle, bu etkileşimli bölgeler, proteinlerin yapısını ve kararlılığını koruyan moleküler ağları temsil eder [ 15 ] .
2 Telafi Edici Epistatik Model
Kondrashov ve ark.’nın [ 16 ] yirmi yıldan uzun bir süre önce yaptığı çalışma, insan dışı memeli türlerindeki vahşi tip alellere karşılık gelen bir dizi insan patojenik varyant örneği sağlamıştır. Bu varsayımsal telafi edilmiş varyantlar, insanlarda hastalıkla ilişkilendirilmiş ancak diğer türlerin genomlarında vahşi tip aleller olarak bulunan varyantları temsil eder. İnsanlar ve diğer memeli türleri arasındaki kısa evrimsel mesafeyi hesaba katarak, çoğu durumda, insanlardaki patojenik bir varyantın herhangi bir diğer memelide aynı zararlı etkiye yol açacağını sezgisel olarak bekleyebiliriz. Ancak, bu durumun bazı varyantlar için geçerli olmadığı, muhtemelen aksi takdirde zararlı bir ikamenin patojenik etkisini geri döndürebilen veya azaltabilen telafi edici ortakların varlığı nedeniyle görülmektedir. Orijinal çalışmalarında, yazarlar, diğer memelilerdeki referans alel olarak çeşitli genlerde insanlarda hastalıkla ilişkili aleller bulmuşlardır. Bunlardan bazıları G6PD , GALT , PMM2 ve PAH gibi doğuştan metabolizma hatalarıyla (IEM’ler) ilişkili genlerdir . Memelilerde yapılan sonraki çalışmalar [ 17 – 21 ] ek örnekler bildirmiştir. Örneğin, rhesus maymunu genomunda [ 18 ], fenilketonüri ile ilişkili fenilalanin hidroksilaz geninde ( PAH ) iki patojenik varyant tanımlanmıştır . Ancak yazarlar fenilalaninin serum seviyelerini test etmiş ve beklendiği gibi herhangi bir artış tespit etmemişlerdir. Rhesus maymunu genomunda fenilalanin seviyesinde herhangi bir artımlı artış olmaksızın PAH’da biallelik varyantların varlığı henüz açıklanamamıştır, ancak yazarlar tarafından dikkate alınan olası açıklamalardan biri telafi edici partnerlerin varlığı olmuştur. Ancak, bu sorunun henüz ele alınmadığını ve sonuç olarak telafi edici bölgelerin tanımlanmadığını belirtmek önemlidir. Ayrıca, belirli varyantların tolere edilebilirliğindeki farklılıkların, etkileşimde bulunan partnerlerin varlığından ziyade türlere özgü metabolik taleplerden etkilenmesi de mümkündür.
Sinir dejenerasyonuyla ilişkili olan α-sinüklein (SNCA)’daki p.Ala53Thr değişiminin durumu da merak uyandırıcıdır. Bu anlamsız varyant ilk olarak Parkinson hastalığından etkilenen ailelerde tanımlanmıştır [ 22 ] ve protein stabilitesinin azalmasına neden olduğu gösterilmiştir [ 23 ]. α-sinükleinin fare doğal tip ortologu, homolog pozisyonda bir treonin içerir ve bu, diğer bölgelerdeki ek telafi edici değişikliklerin 53. pozisyondaki treoninin zararlı etkisini azalttığını düşündürmektedir. Bunu araştırmak için Losasso ve arkadaşları [ 24 ], insan ve fare doğal tip proteinlerinde ve ayrıca Ala53Thr ikamesini taşıyan insan proteininde karşılaştırmalı moleküler dinamik simülasyonları yürütmüştür. Sonuçları, fare proteininin telafi edici bölgelere sahip olduğunu ortaya koymuştur ve moleküler telafinin kesin mekanizması hala belirsiz olsa da çalışma, molekül içi etkileşimlerin SNCA p.Ala53Thr ile ilişkili nörotoksisite etkisini önlediğini göstermiştir.
3 IEM: Ornitin Transkarbamilaz Örneği
Ornitin transkarbamilaz eksikliği, üre döngüsünün en yaygın doğuştan gelen hatasıdır [ 25 , 26 ]. OTC geni X’e bağlı olduğundan, hastalık kadınlarda çok çeşitli fenotipik belirtiler gösterir ve genellikle erkek hastalarda daha şiddetlidir. Hastalığın erken başlangıçlı bir formuyla ilişkili bir varyant (c.374C>T p.(Thr125Met)) ilk kez yaklaşık otuz yıl önce bir erkek hastada bildirilmiştir [ 27 ]. Beklenmedik bir şekilde, bu varyant [ 27 , 28 ] Genom Toplama Veritabanında (gnomAD) [ 29 ], 12 hemizigot bireyi de içeren T aleli (Met 125 ) için 0,0000153’lük bir sıklıkta mevcuttur. Bu varyantın patojenitesini açıklığa kavuşturmak için daha fazla çalışmaya ihtiyaç vardır ve bunlar OTC geninde bildirilen mozaiklik modellerinin değerlendirilmesini içermelidir [ 30 , 31 ]. Bu varyant şu anda ClinVar’da belirsiz öneme sahip olarak sınıflandırılmaktadır [ 32 ].
Şempanze genomu yayınlandığında [ 19 ], şempanze OTC proteininde 125. pozisyonda bir metiyonin bulundu ve o zamandan beri varsayımsal telafi edilmiş varyantın en iyi incelenmiş örneklerinden biri oldu. Met 125’in patojenik olduğu varsayılarak, Suriano ve meslektaşları tarafından gerçekleştirilen in vitro çalışmalarda şempanze genomundaki zararlı etkisini geri döndürebilecek veya telafi edebilecek bir telafi edici partnerin varlığı dikkate alındı [ 33 ]. Yazarlar, bir Thr 135 ile birlikte bulunmadığı sürece, bir Met 125’in varlığının enzimatik aktiviteyi azalttığını gösterdi . Ensembl veritabanındaki mevcut memeli genomlarının kapsamlı bir araştırması [ 34 ], diğer türlerde de bir Met 125 olduğunu ortaya koydu (Şekil 1 ) ve tüm bu türler genomlarında varsayımsal telafi edici Thr 135’i taşıyordu.
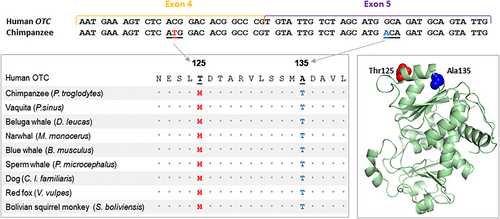
OTC proteini içindeki çiftler arası etkileşim. 125 ve 135. kodonları gösteren ekson 4 ve ekson 5 gen kodlama dizisi. 125. pozisyonda bir Met bulunan Ensembl’den çıkarılan insan ve memeli dizileri arasında kısmi çoklu dizi hizalaması. PyMOL Moleküler
Grafik Sistemi, Sürüm 3.0 Schrödinger LLC ile oluşturulan, doğal insan dizisindeki Thr
125 ve Ala
135 konumlarını gösteren OTC’nin (PDB 1OTH) üç boyutlu yapısı.
Varyantlar arasındaki molekül içi etkileşimin bir diğer boyutu, patojenik ve iyi huylu varyantlar arasındaki etkileşimle ilgilidir. Nötr varyantların in vitro OTC aktivitesindeki rolünü değerlendirmek için Lopes-Marques ve ark. [ 35 ], iki polimorfik iyi huylu varyantın (c.137A>G p.(Lys46Arg) ve c.809A>C p.(Gln270Arg)) farklı alel kombinasyonlarını taşıyan genetik arka planlar oluşturdu. Sonuçlar, her iki polimorfik varyantın da cis’te birleşen minör alellerinin (Arg 46 ve Arg 270 ), vahşi tip arka plana kıyasla OTC aktivitesinde önemli bir artış gösterdiğini gösterdi. Bu mutantlar c.119G>A p.(Arg40His) patojenik varyantıyla birleştirildiğinde, His 40’lı tüm arka planlar enzimatik aktivitede bir azalma gösterdi; ancak bu azalma Arg 270 aleli ile birleştirildiğinde daha az belirgindi ve bu da polimorfik p.Gln270Arg için telafi edici bir rol olduğunu düşündürmektedir. Bu veriler, in vitro bir hücre modelinden elde edilmiş olmalarının getirdiği sınırlamalara rağmen, daha önce nötr polimorfik varyantlar olarak kabul edilenlerin biyolojik rollere sahip olabileceği ve etkilerinin daha fazla incelenmesi gerektiği görüşünü desteklemektedir.
4 Hesaplamalı Patojenite Tahmini Üzerindeki Etki
Önemli bir husus, varsayımsal kompanse edilmiş varyantların patojenite değerlendirmesindeki etkisiyle ilgilidir. Jordan ve ark. [ 36 ], insanlarda mikrosefaliye bağlı BTG2’de (c.421G>A p.(Val141Met)) de novo bir varyantın, çeşitli memelilerde vahşi tip alel olduğunu tespit etti. Yazarlara göre, fonksiyonel analizler patojenitesini göstermesine rağmen hesaplamalı tahminler bu patojenik varyantı iyi huylu olarak puanladı.
Azevedo ve ark. [ 37 ], birkaç memeli türünün proteom dizisini İnsan Gen Mutasyon Veritabanı’nda [ 38 ] açıklanan hastalıkla ilişkili varyantların geniş bir veri kümesiyle çapraz karşılaştırarak bu konuyu ele aldı. Sonuçlar, bu varyantların bir kısmının analiz edilen memelilerden en az birinde vahşi tip alele karşılık geldiğini ortaya koydu. Bu çalışmada [ 37 ], IEM’lerle ilişkili olanlar da dahil olmak üzere en yüksek sayıda varsayımsal telafi edilmiş varyanta sahip olduğu bulunan genler Tablo 1’de gösterilmiştir . Genel olarak, bu varyantlar, insan dışı protein homologlarındaki varlıkları nedeniyle patojenite tahmin araçlarından düşük puanlar gösterme eğilimindedir ve bu da etkilerini değerlendirmek için işlevsel çalışmaların önemini pekiştirir.TABLO 1. Azevedo ve ark. tarafından bildirilen veri setini kullanarak insan hastalığıyla ilişkili genlerde bulunan varsayımsal telafi edilmiş varyantlar [ 37 ]. Kalın yazı tipindeki genler, çevrimiçi olarak erişilebilen Doğuştan Metabolizma Hataları Bilgi Bankası’nda (IEMbase) açıklanmıştır ( https://www.iembase.com , 16 Aralık 2024’te erişilmiştir).
Tazminat Ortaklarının Belirlenmesinde 5 Zorluk
Literatürde tanımlanmış olan varsayımsal olarak telafi edilmiş varyantların sayısız örneğine rağmen, yalnızca birkaç rapor spesifik etkileşimli telafi edici ortağı/ortakları tanımlamayı başarmıştır. İki örnek, yukarıda belirtilen OTC [ 33 ] ve BTG2 [ 36 ] proteinlerini içerir. Başka bir örnek, kan pıhtılaşma Faktörü IX’da [ 39 ] tanımlanmıştır . Özellikle, p.Glu270Lys varyantı, substrat ve inhibitör özgüllüğünü düzenlemek için çok önemli olan 39-döngüde konformasyonel değişiklikler sergiler. Moleküler dinamik analizleri, bazı memeli türlerinin genomunda meydana gelmesi gerçeğine dayanarak, varyantın 271. pozisyonda bir prolin ile eşleştirildiğinde bu yapısal değişikliklerin hafifletilebileceğini ortaya koymuştur.
Akciğer kanseri ve foliküler lenfoma ile ilişkilendirilen poli(ADP-riboz) polimeraz 1’deki (PARP1) p.Val762Ala varyantı ile ilgili olarak başka bir örnek verilmiştir [ 40 ]. Yazarlar, PARP1 p.Ala775Glu varyantının olası bir telafi edici ortak olduğunu tahmin etmek için karşılaştırmalı analizler ve moleküler dinamik simülasyonları kullanmıştır.
Telafi edici varyantlar, belirli patojenik varyantlarla öngörülen birlikte evrimlerine dayanarak tanımlansa da, telafi edici ortakların çeşitli patojenik varyantların bozucu etkisini azaltarak hedef proteinin yapısının stabilize edilmesine de katkıda bulunabilmeleri mümkündür. Bu senaryoda, telafi edici bölgelerin önemi, belirli patojenik varyantlara ortak olma rollerinin ötesine geçmektedir.
6 Sonuç
Burada, insan dışı genomlarda tolere ediliyor gibi görünen insan patojenik varyantlarına genel bir bakış sunarak, molekül içi telafi edici epistatik modelin önemini vurguladık. Önceki verileri tekrar gözden geçirdik ve bu varyantların patojenite değerlendirme araçlarıyla değerlendirildiğinde düşük puanlar alma eğiliminde olduğunu gösterdik. Üre döngüsünün en yaygın doğuştan gelen hatasına odaklanarak, OTC proteinindeki bazı bölgelerin telafi edici bir rol oynayabileceğini gösteren in vitro veriler topladık. Gelecekteki araştırmaların, hastalıkla ilişkili proteinlerde bu olgunun önemini vurgulayacağını ve moleküler etkileşim mekanizmasını ve uyumsuz genotip-fenotip korelasyonlarını açıklamadaki önemini anlamamızda yeni yollar açacağını öngörüyoruz.
Yazar Katkıları
LA, makalenin ilk versiyonunun fikrini ortaya attı ve taslağını hazırladı. LA, TO-M. ve MS-P. verileri toplayıp analiz etti. Tüm yazarlar metnin son haline katkıda bulundu.
Etik Beyanı
Bu makalede yazarlardan herhangi biri tarafından insan veya hayvan denekleri üzerinde yapılmış herhangi bir çalışma bulunmamaktadır.
Onay
Yazarların bildirecekleri bir şey yok.
Çıkar Çatışmaları
Yazarlar herhangi bir çıkar çatışması olmadığını beyan etmektedir.
Makaleye erişmek için lütfen bağlantıya tıklayın: https://onlinelibrary.wiley.com/doi/epdf/10.1002/jimd.70059